A Biosensor Monitoring Approach for Toxic Algae: Field Validation of Selected Probes for Benthic Toxic Species Attached to Macroalgae at Boa Vista Cape Verde Islands
Gerardo Mengs1, Linda K. Medlin2,3*
1EcotoxiLab, Madrid, Spain
2Algae Save the World Consulting, Cardigan, UK
3Department of Electrical Engineering and Electronics, University of Liverpool, Liverpool, UK
*Correspondence to: Linda K. Medlin, PhD, Professor, Department of Electrical Engineering and Electronics, University of Liverpool, Brownlow Hill, Liverpool, L69 3G, UK; Email: linda.crawford@liverpool.ac.uk
DOI: 10.53964/id.2025013
Abstract
Toxic benthic dinoflagellates are predicted to increase with increased temperature stratification and eutrophication. Total rRNA was extracted from cells washed off macroalgae collected at low tide from selected sites on north and western coasts of Boa Vista Cape Verde. The rRNA was tested for the presence of certain toxic dinoflagellates using a molecular barcode to each target species and detected with electrochemical measurement methods in order to establish which macroalga could harbour benthic dinoflagellates. We provide a first rough indication of their density and distribution and probe sensitivity. Toxic planktonic dinoflagellates were also collected from the washes of the macroalgae, indicating that they had likely settled out of the plankton and were trapped on the surface and within the branches of macroalgae. Notably certain benthic dinoflagellates were also found to be abundant in the two plankton samples taken. Electrochemical detection with a biosensor is an alternative cheaper and easier to use method for the detection of toxic algae. A northward progression of emergent toxic tropical species has been now detected with our methods from Cape Verde to the northern Iberian Peninsula because climate change has modified ocean temperatures to enable their survival.
Keywords: Cape Verde, benthic toxic microalgae, probes, electrochemical detection
1 INTRODUCTION
Benthic harmful dinoflagellates, such as species of the genera Gambierdiscus, Ostreopsis, Prorocentrum and Coolia, can produce potent toxins that affect marine organisms the environment and human health and can cause important public health and environmental risks[1]. However, these toxic benthic species, unlike planktonic species, are difficult to monitor using standard whole water samples[2]. Special traps for benthic dinoflagellates have been devised for these microalgae[2]. Benthic species are predicted to increase with increased temperature stratification and eutrophication. Tropical species are spreading northward as climate changes[3].
Some of these toxic benthic dinoflagellates were first reported in the Cape Verde Islands by Soler-Onís et al. [4] whose goal was to make a comprehensive list of microalgae in the Cape Verde archipelago over a wide range of depths and substrates including macroalgae but there was no mention of which macroalgal species were sampled. Here we sampled macroalgae collected at low tide from selected sites on north and western coasts of Boa Vista Cape, Verde washed them and concentrated the cells in the wash. The rRNA extracted from the concentrated cells was tested for the presence of these toxic dinoflagellates using molecular probes. We expand the documentation of certain toxic microalgae illustrating the macroalgae from which they were sampled and the cell numbers of these species dinoflagellates as they were inferred from the molecular probes specific for their detection as a first rough indication of their density, distribution and probe sensitivity. This greatly advances the knowledge of the distribution of these toxic dinoflagellates.
2 MATERIALS AND METHODS
2.1 Sample Preparation
In Boa Vista random macroalgae were collected at low tide but still submerged from 10 sites around the northern and western coasts of Boa Vista in April 2023. Occasionally different macroalgae were collected that were stranded on the beach to broaden the diversity of the macroalgae sampled. The macroalgae represented red, brown, and green macroalgae that were present at each site (Figure 1). These were returned to our portable laboratory where the macroalgae were shaken by hand for 5 minutes in 10L filtered seawater to loosen any unicells that were attached or associated with the macroalgae, either as benthic microalgae or as plankton that had settled on the macroalgae. The 10L were then filtered using the automated environmental sampler with hollow fibre filters (Figure 2) [5]. The first 500mL which we know, from empirical testing, contains 99% of all cells in the sample was filtered onto 3μm nucleopore filters and fixed with Lugol’s iodine. They were returned to CIIMAR, Matosinhos, Portugal where RNA extractions were performed on the filters from the concentrate. No cell counts were made from any of the Boa Vista samples because they could not be quantified at the time of sampling nor were they visually inspected.
|
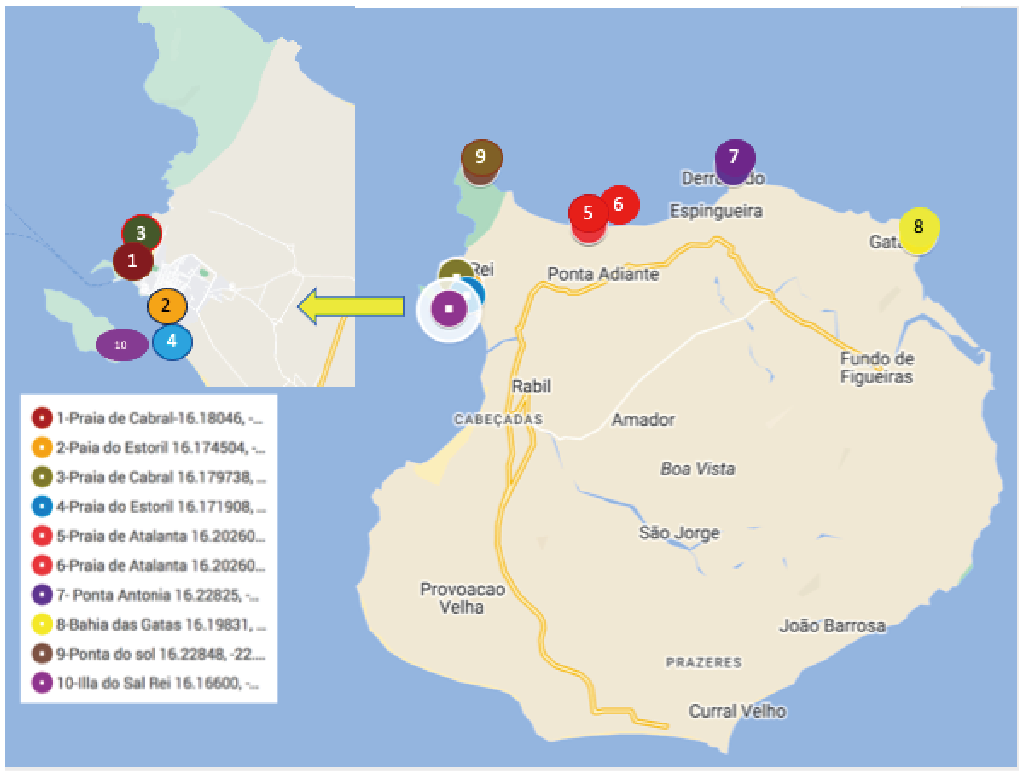
Figure 1. Location of Sampling Sites Around Boa Vista Cape Verde.
|
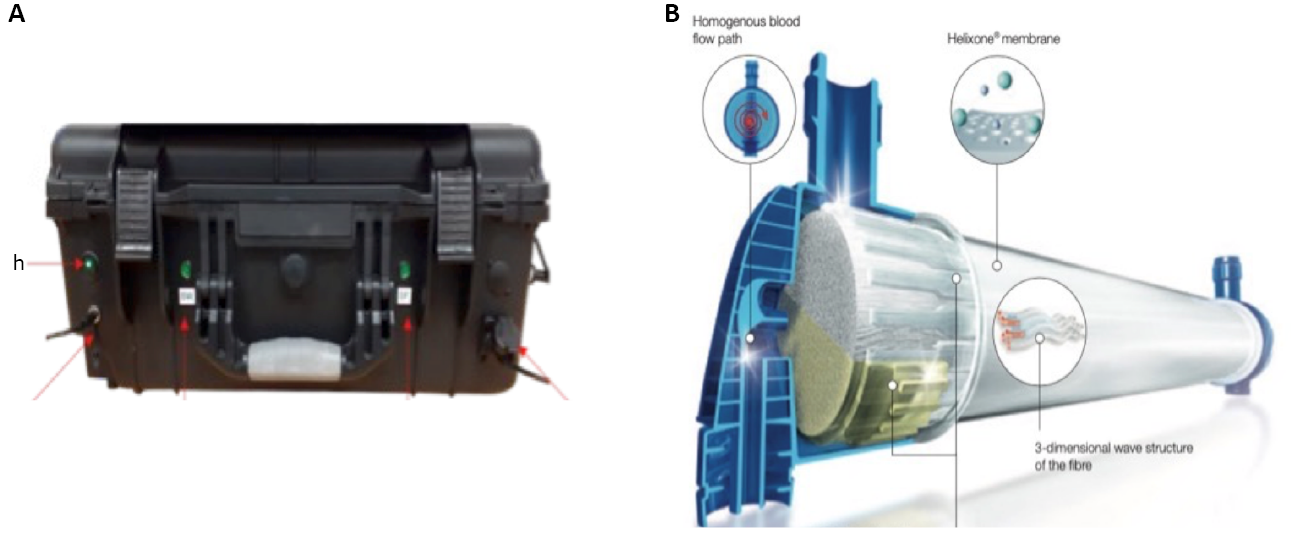
Figure 2. Automated Sample Filtration Unit (A) Manufactured by Q-Technologies Liverpool UK using Hollow Fibre Filters (B) Courtesy of Fresenius.
2.2 RNA Extraction
Total RNA was extracted using TRIzol reagent (Sigma Aldrich Catalogue Number T9424) following the protocol in Lewis et al.[6] DNA and RNA quantities were assessed using a Nano-Drop (ND-1000 UV-Vis spectrophotometer; NanoDrop Technologies) monitoring the absorbance at 260 nm and the 260/280 nm ratio (this ratio varied between 2.11 and 1.39 in all the samples). RNA extractions were very efficient and pure high concentrations of RNA were obtained from each extraction (Table 1).
Table 1. Comparison of the RNA Extraction Concentrations and Their Purity from the 10 Macroalgal Washings
Sample |
Concentration |
A260 |
Ratio |
|
260/280 |
260/230 |
|||
Bv1 |
40.572ng/mL |
1.014 |
1.509 |
Not recordable |
Bv2 |
87.348ng/mL |
2.184 |
1.7 |
0.165 |
Bv3 |
111.573ng/mL |
2.789 |
1.747 |
0.0927 |
Bv4 |
97.947ng/mL |
2.449 |
1.868 |
0.336 |
Bv5 |
191.045ng/mL |
4.776 |
1.853 |
0.561 |
Bv6 |
132.307ng/ mL |
3.308 |
1.773 |
0.425 |
Bv7 |
227.26ng/mL |
5.681 |
1.926 |
0.653 |
Bv8 |
67.533ng/mL |
1.688 |
1.847 |
0.733 |
Bv9/1 |
47.841ng/mL |
1.196 |
1.723 |
0.198 |
Bv9/2 |
38.878ng/mL |
0.972 |
1.795 |
0.0705 |
Bv10/1 |
181.903ng/mL |
4.548 |
1.655 |
0.396 |
Bv10/2 |
184.128ng/mL |
4.603 |
1.655 |
0.41 |
Notes: Washings from site 9 and 10 were so concentrated that the extraction was split into two samples.
2.3 Probe Design
Probes were designed using the ARB program and its probe design function (https://www.arb-silva.de/)[7] and database 119 that was released in 2014. The selected probes were then blasted against Genbank to see if any sequences had been deposited in Genbank since 2014 that matched our probe. If yes, then that probe was discarded and another one designed. The specificity of the probes was tested using a sandwich hybridisation analysis and synthetic target[8]. The two probes involved in the SHA were combined into a single unlabelled probe to use in the field studies with total RNA extraction from the field samples. The rational for changing from the SHA to a single probe for the electrochemical detection is presented in Medlin et al[8].
2.4 Probe Selection
We tested the probes using the single electrode chips on field samples using the newly developed electrochemical method with a single probe. Probes or barcodes from the SSU or LSU rRNA genes to recognise each of these emergent toxic species targeted were designed in EMERTOX (https://cordis.europa.eu/project/id/778069). A total of 50+ probe sets were designed for the sandwich hybridisation method for species apparently spreading into European waters (e.g., Gambierdiscus, Ostreopsis, Coolia species) and a variety of other HAB spp. (e.g., Prorocentrum lima) that already exist in European waters and occur along the northern Iberian Peninsula[9-11]. We used the one probe that combines the first capture and signal probe into one longer probe (C+S) to bind to target RNA for inferring cell numbers from the strength of the electrochemical detection for field tests and monitoring of toxic algae. This probe is unlabelled and electrochemical detection takes place via a labelled antibody[8] and it is this probe that will be commercially developed into an early warning system for toxic algae. The most common use for direct comparisons between samples is to relate the signal for a specific probe with a cell number using a calibration curve Medlin et al[12].
We designed/modified probes or barcodes from the SSU or LSU genes to recognise each of the emergent toxic species targeted in EU EMERTOX. Probes are patent pending but are based on the probes and the methods covered by PCT/DE2003/002124 and PCT/BG2013/051938. A total of 50 probe sets were designed for the sandwich hybridisation method for the species moving into European waters (e.g., Gambierdiscus, Ostreopsis, Coolia and Prorocentrum lima). Each set contains the following probes: capture and signal probes that bind to a synthetic target (Figure 3A) for method 1 used to test for probe specificity [8] and one probe that combines the first capture and signal probe into a single longer probe to bind to target RNA from field material (Figure 3B) = method 2 to infer cell numbers. The specificity test results and the calibration of the probes for quantification are presented in Medlin et al.[8] and Medlin et al[13]. The probes are designed to hybridise between 50-60℃. We use an electrochemical detection with the probes immobilised to magnetic beads on an electrode.
|
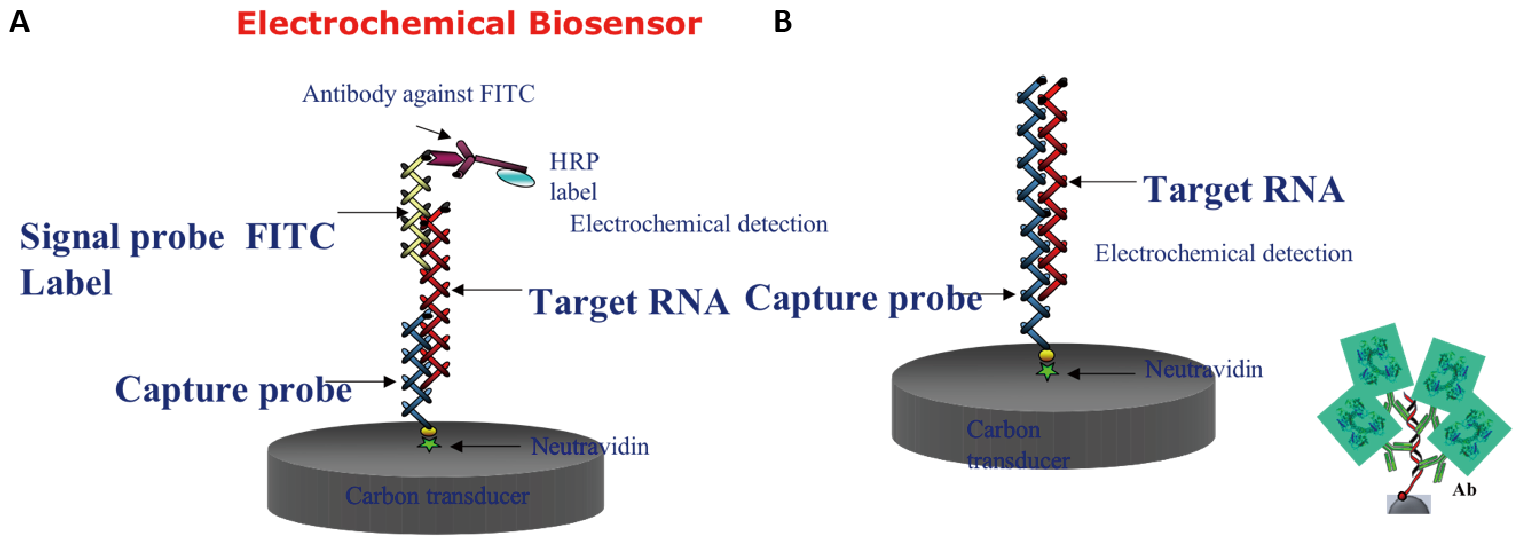
Figure 3. Comparison of Probe Constructs on the Single Electrode Chip. (A) Typical sandwich hybridisation with capture probe immobilised to the electrode and a signal probe with FITC label that initiates the electrochemical reaction when the antibody to FITC is added. (B) Single capture probe with no label immobilised to the electrode that binds to target RNA. Antibody (Ab) recognises the DNA/RNA heteroduplex to initiate the electrochemical reaction.
Twelve probes were selected to test the field samples using single electrode chips either singly or as a pooled group either because of time restrictions or because cell counts cannot distinguish the species and they are group by size (e.g., Pseudo-nitzschia) or by the genus if the species are not counted at all. The probes we tested here include Alexandrium minutum, Alexandrium ostenfeldii, Dinophysis acuta, Dinophysis acuminata, Dinophysis caudata, cells of Pseudo-nitzschia larger than 3μm (mixture of P. seriata, australes, multistriata) cells of Pseudo-nitzschia smaller than 3μm (mixture of P. calliantha, arrenysis, dolorosa) Gymnodinium catenatum, Gambierdiscus three species (mixture of G. eccentricus, australes, silvae) the genus Ostreopsis and a mixture of four Coolia spp. (monotis, tropicalis, canaryensis, and cf. canaryensis).
2.5 Probe Hybridisation and Data Analysis
The hybridisation protocol for electrochemical detection was performed by assembling capture probes for each species to be tested by hybridization to RNA extracted from the concentrated wash of the seaweeds. Probes were synthesized as described in Medlin et al[8]. Five μL of magnetic beads (Strep-MBs 2.8μm Ø 10mg mL−1 Dynabeads M-280 Streptavidin 11206D from Dynal Biotech ASA Oslo Norway) were added to 50μL of B&W buffer (10mM Tris–HCl solution containing 1mM EDTA and 2M NaCl) in each Eppendorf tube. We placed the tubes in a magnetic rack for 2′ removing the B&W buffer and repeating this step once by flicking the tubes several times to resuspend beads before placing them back in the magnetic rack for 2′ with the lid open before removing all of the liquid.
We prepared 0.1mM stock of the capture probe in the B&W buffer with 1μL of 100μM stock and 999μL B&W buffer and added 25μL of probe solution to tubes with MBs incubating them at 37°C 60′ and then washing them twice with PBS buffer allowing each wash to stay in the magnetic rack for 2′ before removing the liquid. They were then resuspended in 25μL BB buffer (PBS solution containing 1% w/v of purified casein Thermo Scientific, Swindon UK) removing all of the liquid in the last rinse. This step immobilised the probes on the magnetic beads.
1000pM of RNA from each concentrated macroalgal wash extraction were prepared. Hybridisation of the single probe to its target RNA was at 55oC for 30’ with agitation. Following hybridization, the tubes were placed on a magnetic rack for 2’ and rinsed twice with PBS before the antibody solution was applied.
An RNA/DNA antibody (AbRNA/DNA clone: D5H6 from Covalab Bron France) was prepared with 200μL AB RNA/DNA (2μg/μL) + 200μL HRP Poly A (1/50 Dilution) (ProtA-poly-HRP40 Antibodies-Online Aachen Germany). Each tube with the hybridized PCR product/probe received 25μL AB/PolyA mixture and was incubated at 37°C for 30′ with agitation and then rinsed twice with PBS buffer for 2′ on the magnetic rack. All beads were resuspended in 25μL of PBS. Hydroquinone (HQ Sigma UK) was prepared with 1.1mg/rxn = 11mg in 1mL PBS buffer and kept in the dark until used. We prepared H2O2; 51μL in 5mL PBS buffer.
For measurements using the amperometer we inserted a new carbon single-electrode chip into the SENSIT-SMART potentiostat from PalmSens (Houten, The Netherlands). We pipetted each probe/AB/PolyA/MB construct in solution onto one working carbon electrode chip (Dropsens (DRP-110 from Methrom-DropSens Birmingham UK)) and added 25μL HQ as a mediator to facilitate the reduction reaction. We applied a current of −0.2V and when the reading of the electrode was stable, we added 25μL H2O2 recording the reading in μAMPs (see Figure 4). The blank was subtracted from each signal to obtain a normalised signal that if greater than the blank was considered to be a positive signal for the probe/species tested. The positive control for each probe (ca. 1000pM RNA) and the signal of the blank were different for each probe tested. Cell numbers were inferred from each sampling site, which consisted of several macroalgal spp. The blank contained no RNA and the positive control was RNA extracted from a culture of the target species. Thus, for each hybridisation the blank and the positive control was different.
|
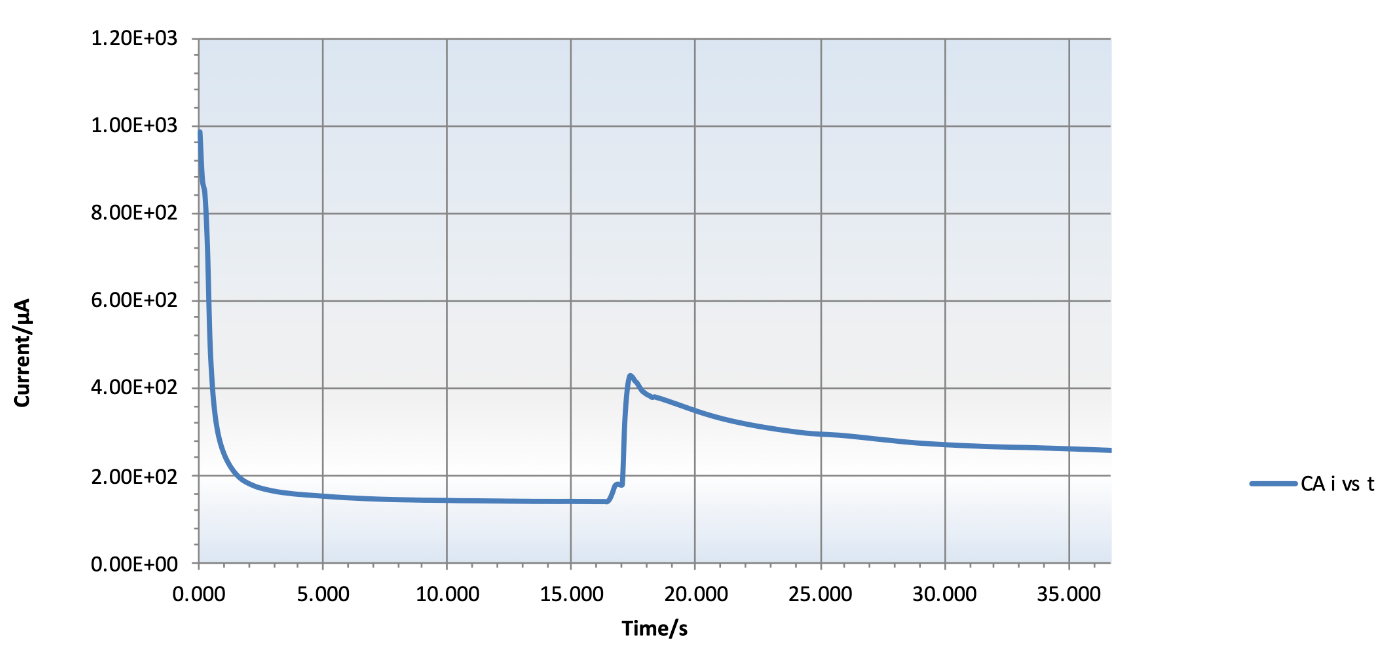
Figure 4. Typical Electrochemical Reaction for 1000pM of RNA Hybridized to the C+S Probe. The rise in the base line is the moment H2O2 is added to the reaction and the signal is proportional to the amount of RNA added to the reaction, which is proportional to the number of cells in the sample.
3 RESULTS
3.1 Macroalgal Species Collected
Photos of the macroalgae collected at each site are shown in Figure S1. One specimen of each type of macroalga was collected at each site.
BV1: This site was located on the north coast of the island and was a rocky tide pool area devoid of a wide variety of macroalgae. One species Enteromorpha sp. was commonly found attached to the rocks.
BV2: This site had no specimens of macroalgae along the tide line and subtidally. The water was very chalky in appearance likely caused by small pieces of coral in the water. Ten litres of water were collected and filtered. It was very difficult to filter because of the suspended sediments.
BV3: This site was similar to site 2 in that no macroalgae were found intertidally or washed up on the shoreline. Again only 10 litres of water was collected and filtered.
BV4: We found mostly brown algae at this site. One species of red algae was collected here.
BV5, BV6, BV7, BV8: These sites were all located along the north coast of Boa Vista and were dominated by 5-6 species of red algae, some brown and green algae, and the occasional colonial blue green alga. See Figure S1 for the photos of each macroalga collected at each site. Morphologies were primarily mixtures of filamentous, blade like or cartilaginous (BV6, BV7, BV8) and cartilaginous (BV5). The macroalgae collected at BV8 were notably void of obvious filamentous epiphytes. BV8 had the most diverse assemblage of macroalgae.
BV9: This site was dominated by three macroalgae of various morphologies.
BV10: This site was an island off the west coast of Boa Vista. We sampled about 10 species of macroalgae attached to the rock surfaces and no samples were taken from subtidal species. No photographs of the macroalgae were made but no new species that those already reported were encountered. RNA extraction of the wash from these macroalgae was difficult because of the high concentration of cells in the wash. We presume that most of the macroalgae were collected from the high intertidal that cells likely could thrive there without being washed away by wave action.
3.2 Hybridisation
Results are shown in Figure 5 for each site and each probe or probe combination as tested in Figure 3B. The signals are high for each probe, but the intensity scale is not the same for each species. This difference reflects the overall strength of the signal of each probe. This, in turn, can influence the inferred cell numbers because of the calibration factor used to convert the normalised signal to cells. The strength of the probe intensity when a given amount of RNA is hybridised can be 1) because each probe is in a different region of the rRNA gene and not all regions have the same accessibility because of the secondary structure of the RNA molecule or 2) because the blank is high. There can also be an interaction between these two factors. For those probes whose intensities reach 400-600μA, these probes are quite strong, whereas those whose maximum signal is less than 250μA are less intense. In two cases, Dinophysis acuta and Gymnodinium catenatum, the blank was quite high (data not shown). All signals were normalised by dividing each probe signal by the blank and if there is a high blank then the signals are reduced and lower cell numbers would be inferred. This effect can be especially seen in those samples where the positive control is zero or very low. This does not mean that the positive control did not give a signal. All positive controls gave a signal but if the blank was too high, then the value for the positive control is reduced or appears as zero and the inferred cell numbers are also lower.
|
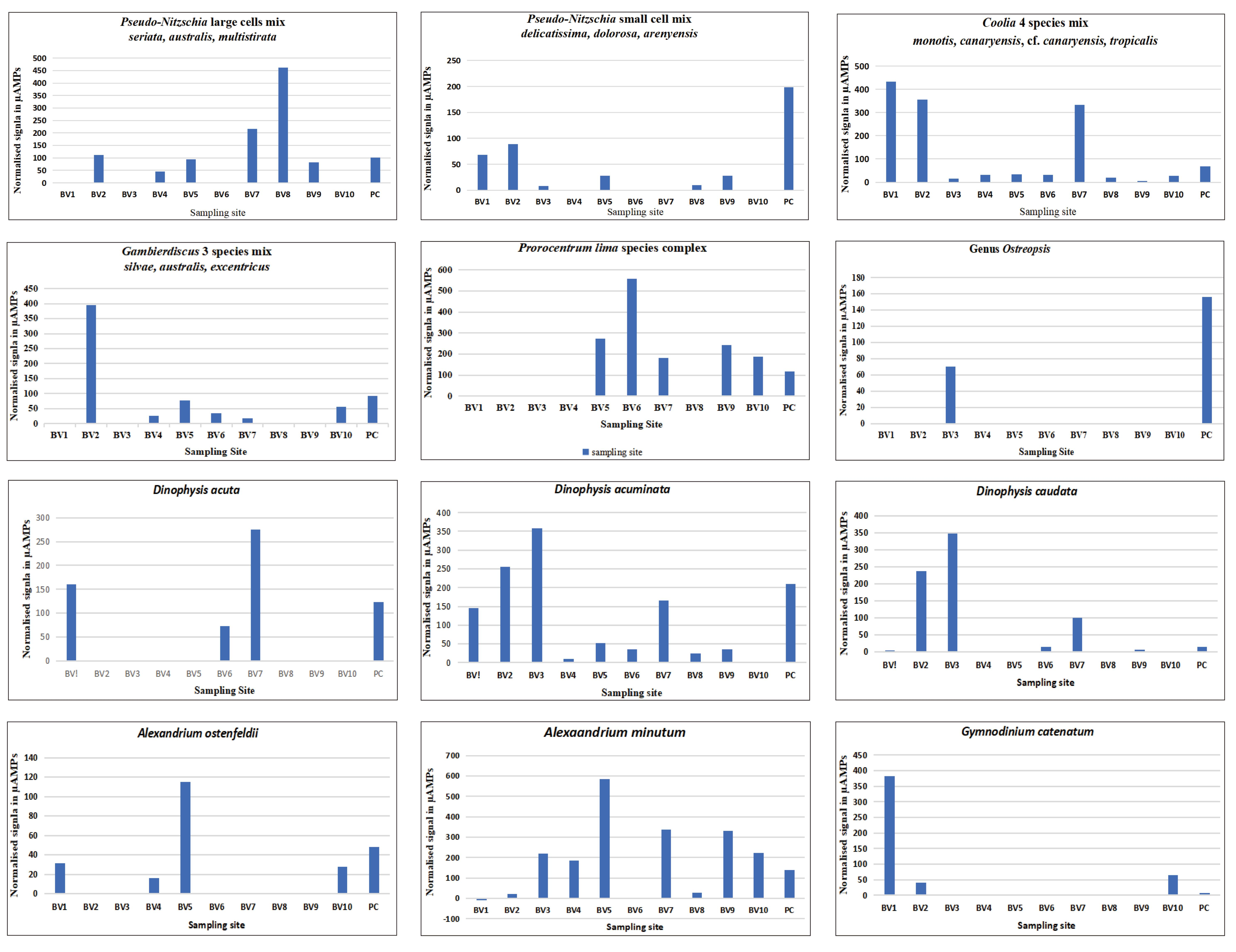
Figure 5. Hybridisation of RNA Extracted from a Wash of the Macroalgae Collected at Ten Sites around Boa Vista. The first eight panels are planktonic species and the next 4 panels are the benthic species.
3.3 Inferred Cell Numbers
The conversion of the biosensor signal to cell numbers is shown in Table 2 for the samples taken in Boa Vista, Cape Verde. We treated all of the samples in the same way in that we collected the seaweeds and shook them manually in the same amount of filtered seawater. There was no attempt to examine the seaweeds for the presence of the toxic algae and thus the inferred cell numbers are only an indication of their concentration on/associated with the seaweeds.
Table 2. Inferred Cell Numbers from the Boa Vista Macroalgal Samples as cells/L
Species |
BV1 |
BV2 |
BV3 |
BV4 |
BV5 |
BV6 |
BV7 |
BV8 |
BV9 |
BV10 |
Pseudo-nitzschia cells>3mm |
0 |
35785 |
0 |
14759 |
30222 |
0 |
69945 |
148905 |
26645 |
0 |
Pseudo-nitzschia cells<3mm |
56979 |
73733 |
6393 |
0 |
23193 |
0 |
0 |
8073 |
23427 |
0 |
Gymnodinium catenatum |
100 |
11 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
17 |
Dinophysis acuta |
99 |
0 |
0 |
0 |
0 |
45 |
170 |
0 |
0 |
0 |
Dinophysis acuminata |
295 |
519 |
731 |
18 |
106 |
73 |
338 |
48 |
70 |
0 |
Dinophysis caudata |
2 |
119 |
174 |
0 |
0 |
7 |
50 |
0 |
3 |
0 |
Alexandrium minutum |
0 |
74 |
723 |
615 |
1932 |
0 |
1113 |
97 |
1097 |
732 |
Alexandrium ostenfeldii |
6316 |
0 |
0 |
3294 |
23331 |
0 |
0 |
0 |
0 |
5680 |
Gambierdiscus (3 spp.) |
0 |
171767 |
0 |
11054 |
33819 |
15120 |
7426 |
0 |
0 |
24859 |
Prorocentrum lima |
0 |
0 |
0 |
0 |
346 |
702 |
230 |
0 |
304 |
236 |
GN Ostreopsis |
0 |
0 |
184 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
Coolia (4 spp.) |
286 |
236 |
10 |
21 |
23 |
22 |
220 |
13 |
4 |
18 |
For the samples from Boa Vista, we found nearly all of the species to be present on the macroalgae at the various sites. No one species was found at all of the sites. Because we collected the algal specimens at low tide it is quite likely that if cells were present in the plankton, then they could have easily been deposited on the macroalgae as the tide retreated. Thus, the chain forming cells of Pseudo-nitzschia and Gymnodinium catenatum could easily be entangled in the mucilage and branches of the macroalgae, whereas Coolia spp., Prorocentrum lima species complex, Ostreopsis and Gambierdiscus would have been attached to macroalgae.
3.4 Planktonic Species
Large cells of Pseudo-nitzschia (mixture of P. seriata, australes, multistriata): This group of species was detected at nearly all of the sties. Notably it was not detected in the high intertidal site BV10. The highest inferred cell numbers were detected at BV8, which was the site with the maximum diversity of macroalgae.
Small cells of Pseudo-nitzschia (mixture of P. calliantha, areneysis, dolorosa): This group of species was also detected with very low signals at over half of the sites.
Gymnodinium catenatum: This species was detected primary at BV1 with lower numbers at two other sites. BV1 had the lowest diversity of macroalgae outside of BV2 & 3, which had no macroalgae.
Dinophysis acuta: The probes used to detect this species is different from the one in Medlin et al.[12] because separate probes were developed that separated acuta from acuminata. This species was not abundant with cell numbers under 200/L and only occurred only on the north shore at the sites BV1, BV6 and BV7.
Dinophysis acuminata: This species was present at all sites other than BV10, which was the high intertidal site on the island off the west coast of Boa Vista.
Dinophysis caudata: This species was highest at BV2 & 3 and relatively low or missing from the other sites. The lowest number of cells/L was inferred with the probes for this species ( 2, 3, 7 cells/L).
Alexandrium minutum: This species was present in nearly all the samples but in relatively low numbers (Table 1).
Alexandrium ostenfeldii: The probe for this species is not as strong as that for A. minutum. It was present in only 4 of the 10 samples but with a maximum of 23331 at BV5 on the north shore.
3.5 Benthic Species
Gambierdiscus three species (mixture of G. eccentricus, australes, silvae): This cluster of species was detected primarily at site BV2 where we did not collect any macroalgae so it was in the water column and not attached to any seaweeds. Minor signals were detected at other sites even at the high intertidal site at BV10.
Prorocentrum lima species complex: was collected primarily on the northwest side of the island with strong signals. It was also collected in the high intertidal site at BV10.
The Genus Ostreopsis: This genus was detected only at one site BV3, which was devoid of macroalgae species.
Four Coolia spp. (monotis, tropicalis, canaryensis, and cf. canaryensisi): This group of species was detected primarily at three sites, with very low signals at the remaining ones. At two of them, Coolia was detected in the wash from the macroalgae, but at BV2 high numbers were found in the plankton because at this site no macroalgae were collected. Figure 6 shows the calibration curves of Coolia canaryensis and cf. canaryensisis, which were performed as detailed in Medlin et al.[13] for the two other species of Coolia. Four species probes were pooled to test for this genus based on their known distribution in the South-eastern Atlantic.
|
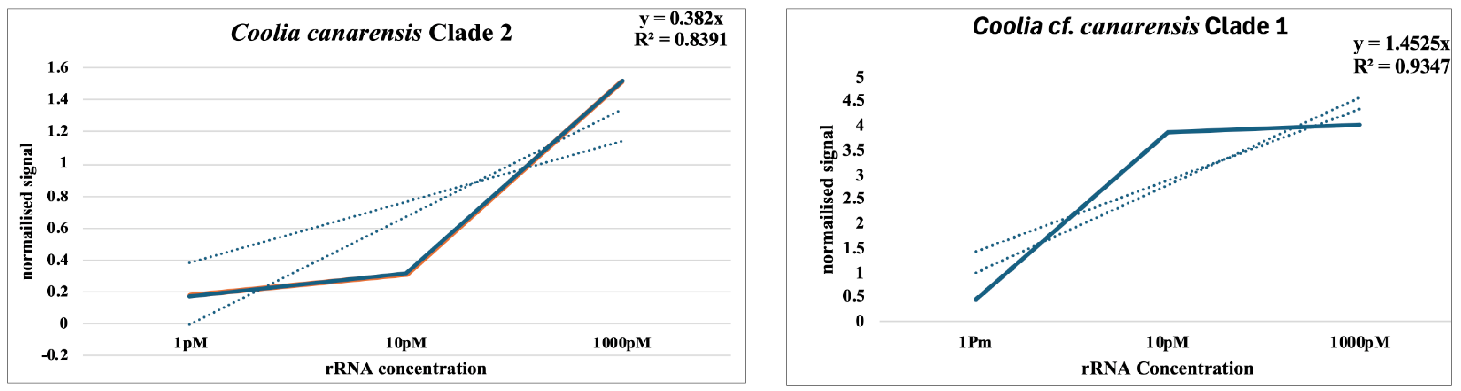
Figure 6. Calibration Curves of the Probes for the Two Clades that Have Been Assigned to Coolia Canaryensis. These slopes were averaged with those of C. monotis and C. tropicalis in Medlin et al.[13] to obtain an average value to infer the cells belonging to this cluster of four Coolia Spp.
Most notable are the signals for Gambierdiscus and Ostreopsis. Gambierdiscus was already known from the Cape Verde Islands[4] but its numbers are quantified here molecular for the first time. Highest numbers were from the plankton samples, and fewer numbers were washed off the various macroalgae. BV4, 5, 6, 7 had only one macroalgal species in common (Dictyota sp.) BV5, with the next highest inferred numbers, had a good mixture of filamentous macroalgae.
4 DISCUSSION
Molecular methods are now widely used in environmental studies. Oligonucleotide probes or barcodes can detect individual species or even strains of species. Our detection of the target species takes place with a barcode to the target species. In electrochemical signal amplification, an antibody coupled to a horseradish-peroxidase (HRP) enzyme binds to either an FITC-labelled signal probe or to a DNA/RNA heteroduplex. HRP electrochemically converts an inactive substrate to an electroactive product that can be detected amperometrically (recorder molecule). The current signal is proportional to the amount of the bound enzyme (and hence to the target concentration in a sample i.e., the number of toxic algal cells). Electrochemical measurement methods present a cheaper and easier alternative to the fluorescent detection used in microarray hybridisation.
In our field studies along the Iberian Peninsula where molecular signals were validated with cell counts from monitoring agencies, our biosensor detected more cells than counted manually by light microscopy and thus the probes can be successfully applied as an early warning system for toxic algae[12]. In that study, we have been able to detect the northward progression of emergent toxic species extending the northern distribution of these tropical species. Our probes were successfully applied to archived samples stored in formalin from the Hardy continuous plankton recorder and we documented the presence of these emergent toxic species as far back as 2014[14]. In this study, we have shown that our probes are highly specific and are very sensitive detecting a minimum of 3 cells. We identified Ostreopsis only from a plankton sample, which might suggest that it prefers macroalgae that live at depth rather than the intertidal ones we were able to collect. It wasn’t present in the high intertidal sample we collected. Highest numbers of Gambierdiscus were also detected in the plankton samples.
The only information available on microalgae in the Cape Verde Islands was published by Silva [15] from material collected on Boa Vista Island in 1948. Sampling macroalgae in Cape Verde by Soler-Onís et al. [4] identified a total of 140 microalgae species. The most numerous group with 108 species were the diatoms. Among the dinoflagellates, Gambierdiscus excentricus, Ostreopsis lenticularis and Ostreopsis cf. ovata were identified. Samples were taken from 32 coastal sites sampled over five of the Cape Verde Islands. None of the associated substates that was associate with these microalgae (sandy and rocky substrates, macroalgal beds and coral beds) were reported. Our study has identified several macroalgae collected at each site and has inferred cell numbers washed off them. This is a major advance over this early study because the presence of these microalgae was corroborated with our analysis of the LSU rDNA D1-D3 and the probes for Gambierdiscus are highly specific with no known targets in Genbank. Ciguatera cases has been reported in Cape Verde previously [16]. Moreover, molecular signals of Gambierdiscus genetic material were detected throughout the NE Atlantic from the northern Iberian Peninsula to the Azores by the Tara Oceans expedition [17] indicating that Gambierdiscus can be far more spread in the North Atlantic than previously thought.
Ramilo et al. [18] performed growth experiments on multiple strains of five Gambierdiscus species. G. australes survived at temperatures as low as 15℃ and exhibited the broadest range of temperature tolerances, hence its global distribution, whereas G. excentricus and G. silvae were better adapted to lower temperatures but could not survive below 17oC. All species who tolerated lower temperatures were kept at low temperatures for up to six months and when returned to 25oC started to grow. They also report the monthly average temperatures in coastal waters of the Canary Islands from 2013 to 2018 to range from 17.26℃ to 18.65℃ in winter and from 23.30℃ to 25.44℃ in summer, which would suggest that the Gambierdiscus spp. in the Canary Islands can tolerate low winter conditions and survive until the summer, where it can bloom [4]. They also report other studies that show G. australes can tolerate temperatures as low as 13℃. Thus, there is a potential for Gambierdiscus to survive in coastal waters along the Iberian coast until warmer summer sea temperatures could initiate growth to cause a toxic event. We recorded Gambierdiscus in the CPR samples taken from a cruise up the Iberian Peninsula in 2014[14] and in water samples. It is likely that these species have migrated from the Cape Verde Islands northward to the Canary Islands and are now along the Iberian Peninsula because climate change has modified ocean temperatures to enable their survival.
5 CONCLUSION AND WIDER APPLICATIONS
The large volume of water sampled by our environmental sampler means that monitoring can be done more frequently and cheaper for better assessment of water quality and GES to enable the detection of rare events and track the species development over time to predict the development of a toxic algal bloom. We have shown here that by using a larger sampling volume, the chances of picking up benthic dinoflagellates increases (see results for BV2 & 3 where only the plankton was sampled.) With our system, any species can be detected, one only needs to design a barcode and test its specificity. Even metabarcoding regions can be used as a probe so undescribed uncultured OTUs can be tracked. eDNA likely retained by our dialysis filter so this can also be detected with our probes. Ideally all of the probes should be on a single platform. such as the microarray in the MIDTAL project [6] so that there is a single negative control or blank with which to normalise all of the signals by the same value. We continue to improve our detection methods and the device with which we measure the signal so that our end goal of an early warning system for toxic algae can be achieved.
Acknowledgements
We thank the staff of CIIMAR for allowing us to work in the Blue Biotechnology, Environment and Health laboratory and the Alfred Wegener Institute for Polar and Marine Research for the loan of filtering equipment. Prof. C.A. Maggs (BV4 & BV5) and Dr. S. Fredericq (all other sites) provided identifications of the macroalgae. This project was funded by EU horizons 2020 Emertox Project grant number 778069.
Conflicts of Interest
The authors declared no conflict of interest.
Data Availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Copyright Permissions
Copyright © 2025 The Author(s). Published by Innovation Forever Publishing Group Limited. This open-access article is licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, sharing, adaptation, distribution, and reproduction in any medium, provided the original work is properly cited.
Author Contribution
Medlin LK and Mengs G collected the field samples. Medlin LK performed the RNA extractions, the hybridizations, the analyses, and wrote the initial version of the manuscript. Mengs G critically read the manuscript and drafted Figure 1.
Abbreviation List
HQ, Hydroquinone
HRP, Horseradish-peroxidase
References
[1] Wakeman KC, Yamaguchi A, Roy MC et al. Morphology, phylogeny and novel chemical compounds from Coolia malayensis (Dinophyceae) from Okinawa, Japan. Harmful Algae, 2015; 44: 8-19.[DOI]
[2] Berdalet E, Tester P A, Chinain M et al. Harmful algal blooms in benthic systems: Recent progress and future research. Oceanography, 2017; 30: 36-45.[DOI]
[3] Wells ML, Trainer VL, Smayda TJ et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae, 2015; 49: 68-93.[DOI]
[4] Soler-Onís E, Fernández-Zabala J, Ramirez AS. First records of Gambierdiscus excentricus and Ostreopsis in the Cape Verde Archipelago (Macronesia Central Eastern Atlantic). Harmful Algae News, 2019; 63: 8-10.
[5] Robla J, García-Hierrro J, Alguacil FJ et al. Determination of the efficiency of filtration of cultures from microalgae and bacteria using hollow fiber filters. Environ Sci-Wat Res Technol, 2021; 7: 1230-1239.[DOI]
[6] Lewis J, Medlin L, Raine R. MIDTAL (Microarrays for the Detection of Toxic Algae): a protocol for a successful microarray hybridisation and analysis. Koeltz: Oberreifenberg, Germany, 2012.
[7] Westram R, Bader K, Prüsse E et al. ARB: a software environment for sequence data. Handbook of molecular microbial ecology I: metagenomics and complementary approaches. Wiley-Blackwell: Hoboken, USA, 2011: 399-406.[DOI]
[8] Medlin LK, Gamella M, Mengs G et al. Advances in the detection of toxic algae using electrochemical biosensors. Biosensors, 2020; 10: 207.[DOI]
[9] David H, Laza-Martínez A, Miguel I et al. Ostreopsis cf. siamensis and Ostreopsis cf. ovata from the Atlantic Iberian Peninsula: Morphological and phylogenetic characterization. Harmful Algae, 2013; 30: 44-55.[DOI]
[10] Seoane S, Siano R. New limits of Ostreopsis distribution in the Bay of Biscay: a first report of Ostreopsis in Santander Bay, Cantabria (Northern Spain). Harmful Algae News, 2018, 60: 10.
[11] Santos M, Oliveira PB, Moita MT et al. Ocurrence of Ostreopsis in two temperate coastal bays (SW iberia): Insights from the plankton. Harmful Algae, 2019; 86: 20-36.[DOI]
[12] Medlin LK, Mengs G, Rodriguez F et al. Detecting emergent toxin-producing microalgae in the Northeast Atlantic: Novel biosensor documents toxin-producing taxa (Gambierdiscus and Ostreopsis) along the Iberian coastline. Phycologia, 2025 (In press).
[13] Medlin LK, García-Portela M, Rossignoli AE et al. A biosensor monitoring approach for toxic algae: Construction of calibration curves to infer cell numbers in field material. Harmful Algae, 2024, 138: 102697.[DOI]
[14] Mengs G, Stern RF, Clarke JL et al. Mapping selected emergent marine toxin-producing organisms using historical samples with two methods (biosensors and real-time PCR): a comparison of resolution. Appl Microbiol, 2024; 4: 312-328.[DOI]
[15] Silva SE. Contribution al'etude du microplankton de dakar et des regions maritimes voisines. Bull Inst Fr Afr Noire Ser A Sci Nat, 1956; 18: 335-371.
[16] Costa PR, Churro C, Rodrigues SM et al. A 15-Year Retrospective Review of Ciguatera in the Madeira Islands (North-East Atlantic, Portugal). Toxins, 2023; 15: 630.[DOI]
[17] McGirr S, Clarke D, Kilcoyne J et al. Insights into the discrepancy between Azadinium spp. and azaspiracid toxins near strategically important aquaculture operations in the west and southwest of Ireland. Estuar Coast Shelf Sci, 2021; 262: 107622.[DOI]
[18] Ramilo I, Figueroa R I, Rayón-Viña F, et al. Temperature-dependent growth and sexuality of the ciguatoxin producer dinoflagellate Gambierdiscus spp. in cultures established from the Canary Islands. Harmful Algae, 2021; 110: 102130.[DOI]
[19] Soler-Onís E, Fernández-Zabala J, Ojeda-Rodriguez A et al. Bloom of Gambierdiscus caribaeus in the temperate-subtropical waters of El Hierro, Canary Islands (north East Atlantic). Harmful Algae News, 2016; 55: 14-17.


