Human Polyomavirus 2 (JC Virus): Understanding Host-pathogen Interactions and Medical Challenges within an Eco-evolutionary Framework
Ramakrishnan Sitaraman1*![]()
1Department of Biotechnology, TERI School of Advanced Studies, New Delhi, India
*Correspondence to: Ramakrishnan Sitaraman, Department of Biotechnology, TERI School of Advanced Studies, 10 Institutional Area, Vasant Kunj, New Delhi 110070, India; Email: minraj@gmail.com
DOI: 10.53964/cme.2026002
Abstract
Human polyomavirus 2 (JC virus) is a neurotropic virus that is ubiquitous in the human population, and most carriers remain asymptomatic for life. However immune suppression, whether natural or because of medication, can lead to virus activation often leading to the fatal condition of progressive multifocal leukoencephalopathy. This critical review provides a survey of the mechanisms of environmental transmission between hosts, the cellular and molecular biology of JC virus-host interactions, and the available therapeutic options – both approved and experimental. It also highlights gaps in our knowledge of the biology and pathogenesis of this virus, and the challenges involved in better understanding the mechanistic basis of JC virus reactivation as well as tailoring therapeutic strategies to different disease contexts. Finally, the available findings are interpreted within an ecological-evolutionary (eco-evo) framework.
Keywords: progressive multifocal leukoencephalopathy, viral persistence, host-pathogen interactions, latency, reactivation, immunomodulation, neurotropic viruses, opportunistic infections
1 INTRODUCTION
The eponymous John Cunningham virus (officially: human polyomavirus 2, henceforth John Cunningham polyoma virus (JCV)) was first discovered in 1971 from a patient who manifested symptoms of progressive multifocal leukoencephalopathy (PML)[1]. This condition is the result of the JCV infection of oligodendrocytes in the white matter of the brain where it replicates in a lytic fashion, culminating in demyelination of neurons with progressively adverse effects on the cognitive and motor skills of the patient. Once commenced, PML is irreversible and fatal in most cases, unless the immune system is able (or aided) to bring the virus under control again.
JCV is a neurotropic virus that remains latent in the kidney[2,3] as well as other cell types and sites such as peripheral blood B lymphocytes[4,5], and most notably, in the brain[6] of immunocompetent individuals. Under conditions of immunosuppression, as in HIV-infected individuals and those undergoing immunosuppression for organ transplants or treatment of autoimmune disorders such as multiple sclerosis (MS), JCV reactivates and transits across the blood-brain barrier to infect brain tissue. The presence of JCV in normal as well as affected individuals may be detected by seroconversion. Its presence in the central nervous system (CNS) can be specifically detected by the polymerase chain reaction (PCR) amplification of JCV DNA sequences from cerebrospinal fluid (CSF). However, given the ubiquity of JCV in the general population, JCV presence in CSF detected by PCR, a highly sensitive method, could also be indicative of contamination with blood during lumbar puncture. Therefore, definitive diagnosis by taking clinical, imaging and virological evidence into account is recommended by the Neuroinfectious Disease Section of the American Academy of Neurology[7]. Briefly, as outlined in these recommendations, the most frequent clinical signs include cognitive, motor, or sensory impairment, either singly or in combination. Specialized magnetic resonance imaging incorporating fluid-attenuated inversion recovery is recommended for the direct observation and estimation of the extent of brain lesions[7,8]. Thus, according to the AAN consensus statement, definitive diagnosis is achieved when PCR positivity of CSF co-occurs with clinical symptoms and positive imaging scans. PCR positivity combined with either clinical symptoms or positive imaging scans is considered ‘probable’. Most authorities consider brain biopsy to be unnecessary except for a minority of cases where the clinical signs or imaging results are inconclusive, and PCR of CSF is negative for JCV[7]. The simultaneous presence of multifocal demyelination, hyperchromatic, enlarged oligodendroglial nuclei, and enlarged bizarre astrocytes with lobulated hyperchromatic nuclei is termed the ‘diagnostic triad’ for PML. The specific presence of JCV may then be confirmed by antibody staining for virions in oligodendrocytes within tissue sections, or by PCR for JCV in brain tissue sections. To summarize, imaging and clinical observation detect multifocal leukoencephalopathy in the brain, while PCR of CSF and immunohistochemistry of biopsies enable causative attribution to JCV invasion of the CNS.
JCV is a non-enveloped double-stranded (circular) DNA virus belonging to the family Polyomaviridae that includes other human and animal viruses such the archetypical simian virus 40 (SV40) and BK virus (BKV). Several reviews of JCV are available that highlight various aspects of its epidemiology, molecular biology, immunology, pathogenesis and PML[9-13]. Therefore, to avoid redundancy, this short commentary seeks to bring into focus some less-studied and emerging facets of JCV biology and pathogenesis to stimulate discussion and further research – basic, translational and clinical – and attempts to situate this instance of host-pathogen interaction with an eco-evolutionary framework.
2 TRANSIT BETWEEN AND WITHIN HOSTS
Up to 70% of the human population is estimated to be infected with JCV, with a wide range of variation likely attributable to a combination of geographic location and sample sizes, and the type and number of subjects surveyed[14-17]. There is sufficient evidence that humans acquire the virus over their lifetime, because seropositivity progressively increases with age[18,19]. The chronic but asymptomatic nature of JCV infection indicates that this virus has established a presence in the human population over a long period of time, as discussed in greater detail in the concluding section. Given that JCV is a widespread pathogen, the incidence of PML is remarkably low as it can be effectively controlled, but not eliminated by immunocompetent individuals. Notably, even while latent, JCV is shed in the urine[20,21], feces[22] and even semen[23] of a subset of infected, but otherwise healthy and asymptomatic individuals. Thus, it is believed that JCV spreads primarily through contaminated water and food in natural settings (see Figure 1). In the light of its presence in semen, specific subtypes seem to be associated with male infertility[24]. It is additionally possible that JCV could be sexually transmitted. In contrast to JCV presence in the above body fluids/excretions, it is almost absent from blood[25] and oropharyngeal fluids[26]. One particularly interesting study of 116 individuals of which half were HIV-infected (i.e., immunosuppressed) and half were healthy controls found that JCV was not detectable in the saliva of any individual. Only two individuals in either group (control and HIV-infected) exhibited JCV-positive throat washings.
|
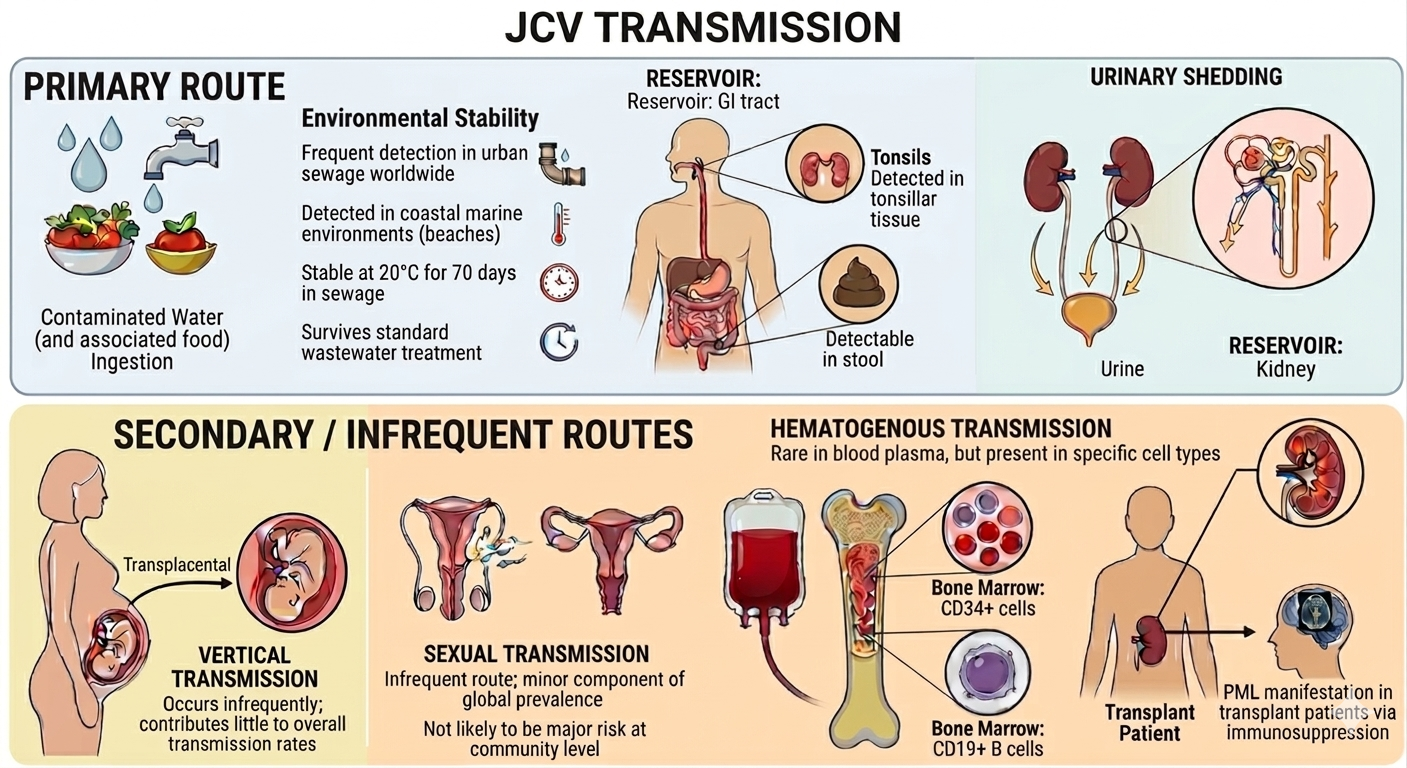
Figure 1. Schematic Overview of Primary and Secondary/Infrequent Routes of JCV Transmission. Image generated using Gemini AI (Google)[27].
In the same study, JCV could be detected in the urine of 70% of all subjects, with the HIV-positive individuals having an approximately 3.4-fold higher median JCV genome copy number compared to the control group[28]. This indicated that JCV was far more likely to be transmitted via urine, as compared to other body fluids. Another surprising finding from the same study was the detection of JCV in the buffy coat (i.e., leukocyte- and platelet-containing fraction of blood) of just one HIV-positive individual. More details regarding the strength of the evidence for various routes of transmission are given in Table 1. The presence of JCV in the buffy coat raises the question as to whether reactivated JCV reaches the CNS by residing within leukocytes (e.g., B cells) or by a different but not mutually exclusive mechanism. The major attachment receptor for JCV is lactoseries tetrasaccharide c (LSTc), a pentasaccharide having a terminal a2,6-linked sialic acid. However, note that the presence of type 2 serotonin (5-hydroxytryptamine) receptors (5-HT2Rs) is required for virus entry into target cells[29,30].
Table 1. Transmission Routes of JCV Ranked in Order of Importance (Primary and Secondary/Infrequent)
Transmission Route |
Supporting Evidence |
Remarks |
Primary Fecal-oral transmission / Urinary shedding |
JCV DNA is detectable in the stool[22], gastrointestinal tracts[31] and tonsillar tissues[5] of healthy adults. It is detectable in urban sewage worldwide[32-35] frequently detected in urban sewage worldwide. JCV can be detected even in costal marine environments (beaches) otherwise considered suitable for recreation by virtue of low coliform bacterial counts[36]. JCV is also shed in urine of both healthy and diseased individuals[20,21,28,37,38]. |
Contaminated water (and by association, food) is considered the primary mode of JCV transmission. The gastro-intestinal tract is considered the main reservoir of JCV. JCV has been found to be stable at 20oC for 70 days in sewage[38] and can survive standard wastewater treatment processes[33]. |
Secondary / infrequent Vertical (Mother-to-Child)
Sexual Transmission
Hematogenous transmission (Blood/Organ donation) |
Vertical transmission is an infrequent occurrence and contributes very little to overall rates of transmission[15,39].
JCV DNA has been detected in semen[23,24] and vaginal secretions[40]. .
While JCV is infrequently detected in blood plasma, particular cell types in the blood may host the virus[4,25,28,41]. In one study, patients undergoing immunosuppressive treatment for MS were found to harbour JCV within CD34+ hematopoietic stem cells and CD19+ B cells within the bone marrow[42].
There are cases of transplant patients manifesting PML due to immunosuppressive drugs administered as part of the transplantation protocol. Kidney transplant patients may be particularly vulnerable, given that the kidney is the main reservoir of JCV[43-47]. |
While proven possible, this is considered an infrequent route compared to horizontal transmission after birth.
Most researchers view this as a secondary or minor route rather than a primary driver of the virus's high global prevalence.
Not likely to be a major transmission risk at the community level. |
This receptor combination was found to be expressed in the kidney (the best-known reservoir of JCV within the body) and the choroid plexuses of the brain, but LSTc is absent from brain parenchyma (where 5-HT2Rs are expressed)[48], which is the precise region of the brain where JCV causes the most damage.
Recent research employing model cell culture systems implicates JCV-containing, host-derived extracellular vesicles as an alternative, receptor-independent mechanism of invasion of target cells by JCV within the brain[49-51]. Extracellular vesicles are classified into subtypes based on their origin and surface markers: Exosomes originate from the endosomal pathway and typically possess tetraspanin proteins. Microvesicles and apoptotic bodies originate from the plasma membrane; the former possess integrins, selectins and the CD40 ligand, whereas the latter possess histones[52]. JCV has been detected in blood plasma-borne extracellular vesicles as well[53], which could be a potential, but unproven alternative route of dispersal of the virus originating from other body sites to the brain. Research using cell culture systems indicates that the vesicles containing JCV possess tetraspanins and that their production can be blocked using drugs targeting exosome biogenesis pathways[54] and cellular trafficking[55,56], indicating their origin within the endoplasmic reticulum, rather than the plasma membrane. Drugs targeting 5-HT2Rs[57,58] have also been found to limit viral spread in in vitro studies. More encouragingly, a limited number of case studies indicate that currently available 5-HT2R antagonists such as mirtazapine[44,59-64] that are commonly used as anti-depressives and anti-psychotics can significantly retard the progression of multifocal leukoencephalopathy either on their own, or as adjuncts to immune reconstitution. More extensive and systematic studies are required to evaluate the advisability of repurposing 5-HT2R antagonists along with immune reconstitution in PML patients. While the relative importance of receptor-mediated and -independent modes of JCV entry into target cells is unknown, the further development of inhibitors of EV production and intracellular trafficking could potentially add to the treatment options currently available.
|
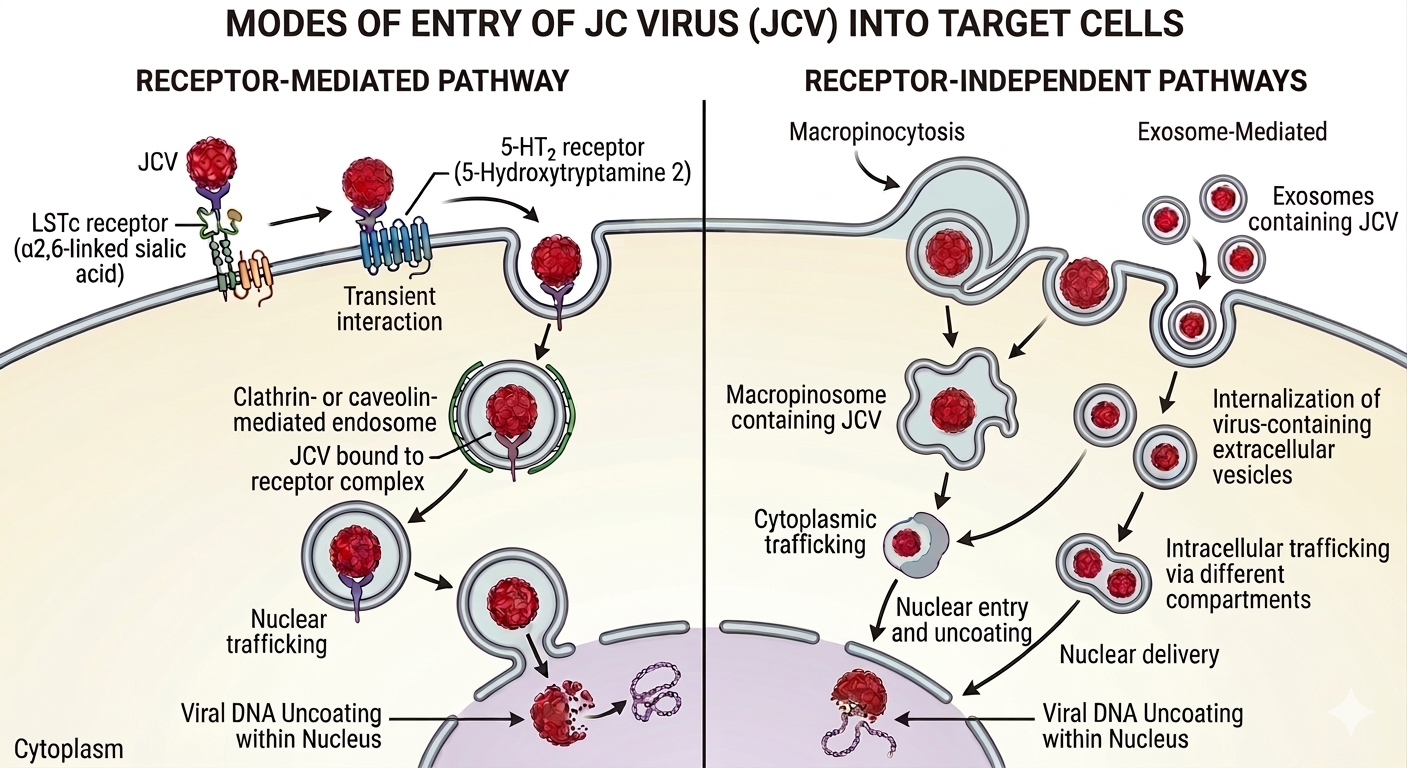
Figure 2. Receptor-mediated (left) and Receptor-independent (right) Pathways of JCV Entry into Target Cells. The receptor mediated pathway involves binding to LSTc receptor followed by transient binding to 5-HT2 receptors, that eventually results in internalization via the endosomal pathway (left). The receptor- independent pathway involves micropinocytosis or fusion exosomes containing JCV with the plasma membrane (right). Image generated using Gemini AI (Google)[27].
3 RESTORING IMMUNE FUNCTION AND BALANCE TO RESOLVE PML
Immunosuppression can result naturally from various types of immunodeficiency diseases, particularly HIV infection, or be deliberately induced during organ transplantation, and during the treatment of autoimmune diseases and different types of cancer. Currently, no JCV-specific, approved antivirals or other therapies are available. The only way to protect a patient exhibiting signs of JCV reactivation is to attempt immune restoration/reconstitution (perhaps with adjuncts, as described in the previous section) and thereby limit viral multiplication and spreading[65]. In the case of HIV-infected patients, the most direct method of immune reconstitution is to administer highly active anti-retroviral therapy (HAART), more recently termed combined anti-retroviral therapy (cART). Unfortunately, it has been observed that HAART in particular, and immune reconstitution in general, can lead to a rebound that manifests in a hyperinflammatory state termed ‘immune reconstitution inflammatory syndrome’ (IRIS). This is accompanied by emergence of PML (termed ‘unmasking’) as a result of a restored immune response against JCV that damages neural cells harboring the virus[66]. Worse, even immune reconstitution may not be effective in stopping the progress of PML in some cases[67,68], which points to rare, individual specific factors (e.g. age, sex, genetics and environment) that may influence the final outcomes. However, more directed efforts at immune restoration may also be warranted. Most notably, pembrolizumab – a humanized antibody against the programmed cell death protein 1 (PD-1) used in cancer immunotherapy – has produced mixed results in the case of PML[69-72]. The reasoning behind its usage is that high expression levels of PD-1 on the surface of CD4+ and CD8+ lead to apoptosis; therefore, blocking PD-1 (or its ligands PD-L1 and -L2) should lead to improved immune responses that would eliminate JCV. The reasons for the observed variation are unknown, and may possibly reflect unknown genetic, physiological or environmental factors. An overview of various therapeutic strategies and agents/protocols for JCV-associated PML with salient details is provided in Table 2[72-79].
Table 2. Overview of Therapeutic Strategies and Therapeutic Agents/Protocols Used in JCV-associated PML
Therapeutic strategy |
Therapeutic agent/protocol |
Remarks |
Immune restoration/ reconstitution |
Cessation of immunosuppressive treatment
Intravenous immunoglobulin
HAART/cART in HIV patients |
Only one recent case report available of an HIV patient’s PML due to JCV being controlled[73]; further validation required.
Effective, but attendant risk of IRIS and unmasking of PML (see text). |
Directed immuno-modulation |
Adoptive transfer of JCV-specific T lymphocytes
Immune checkpoint inhibitors (ICIs) |
A few case reports of successful outcomes are available[74-76]. Considered one of the most effective among upcoming treatment methods.
A recent retrospective, non-controlled analysis of outcomes of therapy targeting PD-1 and/or its ligands (PD-L1 & -L2) indicated that slightly more than half of patients (41/79) survived for more than one year. However, the study also found adverse events in 24 out of 79 patients, suggesting that genetics, pre-existing conditions and environmental factors have to be taken into account[72]. Pre-existing antiviral immunity enhances effectiveness of ICIs[77]. |
Antivirals |
Tenofovir alafenamide fumarate
Other antivirals |
Repurposed broad-activity antiviral, already approved for use against HIV & hepatitis B virus. Achieved reduction of JCV titres to undetectable levels in a single case study only[78].
A comprehensive listing of antivirals targeting multiple steps of JCV life cycle is provided in Table 1 of Kaiserman et al.[79] |
Notes: The strategies have been classified as immune reconstitution, directed immunomodulation and antivirals. Therapeutic agents have been grouped into types, rather than listing individual ones for reasons of brevity and to avoid redundancy. At present, there is no JCV-specific approved treatment or vaccine.
At this point, it is important to note that there are additional wrinkles to be ironed out in this somewhat general account of immune restoration. On occasion, the immune system may fail to keep JCV under check even in the background of general hyperinflammation, as in sarcoidosis. Sarcoidosis is an inflammatory disease of unknown etiology wherein granulomas are spontaneously formed in multiple organ systems, particularly in the lungs and lung hilar lymph nodes (bilateral hilar lymphadenopathy). It is accompanied by a disequilibrium between T cell subsets that manifests as anergy (i.e., failure to respond to antigen stimulation) in spite of intense inflammation[80]. Extrapulmonary sarcoidosis presentation varies with age, sex and ethnicity[81,82]. It is interesting (and confusing) to note that JCV reactivation and consequent PML may develop not only in patients who are administered immunosuppressants, but also in patients not receiving such therapy[83-85]. Specifically, increased T regulatory (Treg) cell frequency is accompanied by a decrease in CD4+ T cell numbers. This condition can be rescued by administering infliximab – a chimeric anti-TNF-a monoclonal antibody[84]. This is because TNF-a promotes the expansion and proliferation of Tregs[86] that, in turn, exert an antiproliferative effect on CD4+ T cells[80]. This brief excursus into JCV reactivation and PML in the context of sarcoidosis strongly suggests that future therapeutic strategies against JCV would benefit from a detailed understanding of how the equilibrium and interactions between populations of diverse T cell subsets impacts both health and disease.
4 AN ECO-EVOLUTIONARY PERSPECTIVE OF JCV INFECTION
No biological entity is an island. From the viewpoint of two different interacting organisms, the range of outcomes for each of them can depend on the abiotic environment (e.g., survival under given conditions of temperature or humidity), their physiological state (e.g., nutritional status of the host, virulence of the pathogen), the specific biological context of the interaction (e.g., historical contingency arising from pre-existing interactions with other organisms) and the mode(s) of transmission available (e.g. insect vectors, fomites, water). It is becoming increasingly clear that either partner in a symbiotic association may move along this continuum of association types over time[87], and that organisms may adopt different strategies in a context-dependent manner to secure their fitness. A particular association may become long-lasting due to mutual interdependence for ensuring the biological fitness of either partner (nitrogen-fixing bacteria in leguminous plants) or due to the ability of one of the partners to irreversibly ‘enslave and exploit’ the other (as in the case of mitochondria and chloroplasts within eukaryotic cells) or due to one of the partner’s ability to parasitize the other in spite of the host’s efforts at defending itself or the host’s ability to tolerate some level of persistence. Therefore, from the viewpoint of the pathogen, reducing virulence to enable persistence is a fitness-enhancing strategy. This may be contrasted with the case of pulmonary anthrax infection by Bacillus anthracis, in which the high fatality rate of the hosts (humans or ruminants) does not present an obstacle to transmission for the bacterium, as it is able to persist as a highly resistant and durable spore for extended periods of time in the soil.
Another important issue to be considered is the difference in the generation times of the participating organisms, as it is crucial to determining evolutionary trajectories for each of them. In the classic case of bacteria and bacteriophage, there seems to be a perpetual ‘arms race’ between the host (bacteria) and the pathogen (phage) where selection imposed by phage infection results in the evolution of bacterial defenses (both specific and non-specific) to eliminate phages which, in turn, amounts to imposing selection on the phage populations for variants that are able to evade or resist or are otherwise unaffected by bacterial defenses. Both organisms reproduce rapidly and attain large population sizes. This may be contrasted with humans and microbial pathogens which is germane to our context, where there is a major gulf in terms of both population size and generation time resulting different constraints on the interacting organisms that influence the ensuing outcomes. We parenthetically note that human populations have be generally low for the majority of the duration that our species has inhabited the earth, with major increases in population having occurred relatively recently as a result of profound changes in our habits and habitat caused firstly by the agricultural revolution (~8th millennium B.C.) and secondly, the industrial revolution (~18th century A.D.). As a result, the ecological context and evolutionary constraints have changed with unprecedented rapidity, particularly with the advent of large-scale transcontinental travel, as the world discovered to its collective cost during the SARS-CoV-2 pandemic that originated in Wuhan, China. This background is necessary to keep in mind while formulating public health policy, even as research continues on individual-focused preventive and therapeutic strategies for various diseases.
Moving from populations to individuals, we note that in host-pathogen interactions, a vigorous and effective immune response need not necessarily culminate in a favorable outcome for the host. It has been proposed that the cost and benefit of immune responses may be understood within a ‘damage response framework’[88]. In this view, increasing immune responses to a pathogen do not necessarily result in increased benefits for the host. Both specific and non-specific immune responses (e.g. cytolysis of infected cells and production of reactive oxygen species respectively) can damage host tissues as well. Therefore, the graph of the costs and benefits to the host plotted against the strength of the immune response follows a parabolic U-shaped curve. Essentially, a weak immune response results in major (adverse) costs to the host, that could culminate in death. As the immune response becomes stronger, these costs to the host are reduced as the pathogen is either eliminated altogether, or evolves to a state of latency. But notably, a stronger immune response beyond this point starts affecting the host adversely due to tissue damage and may result in death, without necessarily eliminating the pathogen. The occurrence of IRIS in instances of immune restoration[46,89] indicates that the mounting of a balanced immune response is perhaps as critical as the mounting of an immune response itself.
Finally, in devising preventive and therapeutic strategies against microbial diseases in humans and animals, the innate and adaptive immune systems have been (rightly) viewed as being critical to the desired outcome, which is resistance to the pathogen. However, this neglects the phenomenon of host tolerance to pathogens that may be significant and has been known in plants for a long time[90,91], but is less well-studied in animals and humans[92]. In tolerance, the host responds to colonization by pathogens with physiological adjustments, and the pathogen itself may evolve towards a state of attenuated virulence[93]. As a result, significant pathogen populations persist in an asymptomatic host, without any observable deleterious effects.
In the specific case of JCV, it is to be noted that its chronic persistence is not known to cause any kind of pathology or symptoms in the majority of immunocompetent individuals. Given that humans are the only natural reservoir for JCV and its mode of transmission is likely through the orofecal route or through airborne droplet infection, the survivability of JCV in the environment is probably low (in contrast to B. anthracis discussed above). Note that JCV is detectable in sewage[32,33,35,94-99] and stable in it for up to 70 days at 20oC[38]. Recent studies of JCV seroconversion rates during the COVID-19 lockdowns at seem to indicate that airborne droplet infection is likely a minor or insignificant contribution to JCV transmission[100,101]. Furthermore in a minority of cases, JCV can also be transmitted vertically by human females to their offspring[39]. Based on the discussion in the preceding paragraphs, this could also be interpreted as JCV evolving towards attenuated virulence and persistence in the host which, given the decades-long life expectancy of the host, offers innumerable opportunities for multiplication and transmission.
The outcome of JCV infection in the context of pre-existing diseases/pathogens or co-infecting pathogens, particularly in vulnerable populations such as the elderly and children, is also a matter of concern. At a conceptual level, prior or co-existing disease conditions could potentially bias the immune response such that reactivation of JCV takes place. In other words, changes in selection pressures arising from altered states of humoral and cell-mediated immunity could result in the reactivation of the virus and emergence of neurotropic variants leading to PML or even cause disease in an atypical manner. As readers will recall, JCV-induced PML emerged as a cause of mortality following the global emergence and prevalence of AIDS[102]. More recently, the possibility of PML in multiple myeloma patients has been recognized[103]. Another worrying instance of collateral damage due to JCV concerns COVID-19 patients. Given the general immune dysregulation observed in SARS-CoV-infection, superinfection with SARS-CoV-2 has been observed to exacerbate PML symptoms in multiple myeloma patients[104] and, in one case, triggered PML in an asymptomatic, undiagnosed HIV patient[105]. The hyperinflammatory state induced in COVID-19 patients could result in unexpected adverse consequences of JCV activation at sites other the CNS as well. Likewise, recall the foregoing discussion on PML associated with the inflammatory disease sarcoidosis. In other words, changes in selection pressures arising from altered states of humoral and cell-mediated immunity could result in the reactivation of the virus and emergence of neurotropic variants leading to PML or even cause disease in an atypical manner. For example, Lv and Liu have reported an atypical case of hemorrhagic cystitis (a urinary tract disease) due to JCV in a SARS-CoV-2-infected individual who had received antiviral treatment[106].
Changing selection pressures in terms of altered immune states may also be responsible for the emergence of neurotropic JCV mutants in an instance of intra-host evolution. The viruses with ‘archetype’ sequences in the non-coding control region (NCCR) of the genome conventionally denoted as ‘ABCDEF’ are not neurotropic. However, deletions and partial duplications in the NCCR (e.g. ACEACEF in the mad1 strain and ABCEBCEF in the mad7 strain) render the virus virulent and neurotropic, resulting in PML[107,108]. These variants likely have a competitive advantage over viruses with archetype sequences in the absence of a ‘normal’ immune response[109]. How environmental and physiological triggers influence mutation rates of JCV variants and their differential survival remains to be understood.
In closing, we would like to draw our readers’ attention to an intriguing inverse relationship between the occurrence of JCV viruria and diabetic kidney disease in African-American subjects[110]. Whether JCV is merely a biomarker for some unidentified aspect of normal physiology or whether it has a causal effect on kidney function remains to be investigated. The lack of an unambiguous small animal model is a major limitation in the field. No mouse-adapted strains of JCV are available. However, some success has been achieved through studies in chimeric mice harboring human glial cells[111] and in humanized mice into which human thymus and lymphocytes have been grafted[112]. These and other model systems have been reviewed in detail elsewhere[113]. In spite of these limitations, is amply clear that studies of JCV will not only lead to the progressive refinement of treatment and prevention strategies but also inform our understanding of how chronic pathogens interact with the immune system of the host – when remaining latent as well as when turning rogue.
5 CONCLUSION
JCV infections are widespread, and the virus can persist without incident through the lifetime of immunocompetent hosts. However, in the event of immunosuppression by natural or artificial means, the latent infection is reactivated and causes severe, irreversible brain damage wherein the white matter is progressively destroyed. JCV reactivation is also implicated in oncogenesis. There is much to be learnt about the basic biology of JCV-host interactions and its implications for the management of disease, requiring sustained research into basic, translational and clinical aspects of infection by this ubiquitous virus. Viewing this widespread, chronic yet infrequently pathogenic virus infection within an ecological-evolutionary framework could offer us additional insights into disease management in a public health context.
Acknowledgements
I would like to thank Prof. Prof. Hai-Feng Pan, Chief Editor, Clinical and Molecular Epidemiology, for expediting the review process. My son Sudharman Sitaraman initially drew my attention to JC virus. This paper is dedicated to my parents, Mr. G. Sitaraman and Mrs. Indu Bala for unconditionally supporting my studies.
Conflicts of Interest
The author affirms that he does not have any conflicts of interest.
Disclaimer
This article does not constitute medical advice. No guarantee is expressed or implied regarding the veracity and medical utility of the information provided on external websites and sources. The opinions expressed herein do not represent the views of the TERI School of Advanced Studies or TERI.
Data Availability
All data generated or analyzed during this study are included in this article or are available in the cited references.
Copyright Permissions
Copyright © 2026 The Author(s). Published by Innovation Forever Publishing Group Limited. This open-access article is licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, sharing, adaptation, distribution, and reproduction in any medium, provided the original work is properly cited.
Author Contribution
Sitaraman R contributed to the conceptualization, data curation, investigation, writing – original draft, writing – review & editing.
Abbreviation List
5-HT2R, Type 2 serotonin (5-hydroxytryptamine) receptor
cART, Combined anti-retroviral therapy
CNS, Central nervous system
CSF, Cerebrospinal fluid
HAART, Highly active anti-retroviral therapy
ICIs, Immune checkpoint inhibitors
IRIS, Immune reconstitution inflammatory syndrome
JCV, John Cunningham polyoma virus
LSTc, Lactoseries tetrasaccharide c
MS, Multiple sclerosis
NCCR, Non-coding control region
PCR, Polymerase chain reaction
PML, Progressive multifocal leukoencephalopathy
Treg, T regulatory
References
[1] Padgett BL, Walker DL, ZuRhein GM et al. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet, 1971; 1: 1257-1260.[DOI]
[2] Chesters PM, Heritage J, McCance DJ. Persistence of DNA sequences of BK virus and JC virus in normal human tissues and in diseased tissues. J Infect Dis, 1983; 147: 676-684.[DOI]
[3] Randhawa P, Shapiro R, Vats A. Quantitation of DNA of polyomaviruses BK and JC in human kidneys. J Infect Dis, 2005; 192: 504-509.[DOI]
[4] Lafon ME, Dutronc H, Dubois V et al. JC virus remains latent in peripheral blood B lymphocytes but replicates actively in urine from AIDS patients. J Infect Dis, 1998; 177: 1502-1505.[DOI]
[5] Monaco MC, Atwood WJ, Gravell M et al. JC virus infection of hematopoietic progenitor cells, primary B lymphocytes, and tonsillar stromal cells: implications for viral latency. J Virol, 1996; 70: 7004-7012.[DOI]
[6] White FA, Ishaq M, Stoner GL et al. JC virus DNA is present in many human brain samples from patients without progressive multifocal leukoencephalopathy. J Virol, 1992; 66: 5726-5734.[DOI]
[7] Berger JR, Aksamit AJ, Clifford DB et al. PML diagnostic criteria: consensus statement from the AAN Neuroinfectious Disease Section. Neurology, 2013; 80: 1430-1438.[DOI]
[8] Mori K, Kurokawa M, Harada M et al. Overview of MRI findings in progressive multifocal leukoencephalopathy. Jpn J Radiol, 2025; 43: 1908-1925.[DOI]
[9] Schweitzer F, Laurent S, Cortese I et al. Progressive Multifocal Leukoencephalopathy: Pathogenesis, Diagnostic Tools, and Potential Biomarkers of Response to Therapy. Neurology, 2023; 101: 700-713.[DOI]
[10] Butic AB, Spencer SA, Shaheen SK et al. Polyomavirus Wakes Up and Chooses Neurovirulence. Viruses, 2023; 15: 2112.[DOI]
[11] Atkinson AL, Atwood WJ. Fifty Years of JC Polyomavirus: A Brief Overview and Remaining Questions. Viruses, 2020; 12: 969.[DOI]
[12] Harypursat V, Zhou Y, Tang S et al. JC Polyomavirus, progressive multifocal leukoencephalopathy and immune reconstitution inflammatory syndrome: a review. AIDS Res Ther, 2020; 17: 1.[DOI]
[13] Del Valle L, Piña-Oviedo S. Human Polyomavirus JCPyV and Its Role in Progressive Multifocal Leukoencephalopathy and Oncogenesis. Front Oncol, 2019; 9: 711.[DOI]
[14] Padgett BL, Walker DL. Prevalence of antibodies in human sera against JC virus, an isolate from a case of progressive multifocal leukoencephalopathy. J Infect Dis, 1973; 127: 467-470.[DOI]
[15] Stolt A, Sasnauskas K, Koskela P et al. Seroepidemiology of the human polyomaviruses. J Gen Virol, 2003; 84: 1499-1504.[DOI]
[16] Kean JM, Rao S, Wang M et al. Seroepidemiology of human polyomaviruses. PLoS Pathog, 2009; 5: e1000363.[DOI]
[17] Karimi Dehcheshmeh L, Makvandi M, Timori A. Prevalence of Human Polyomavirus JC and BK in Normal Population. Asian Pac J Cancer Prev, 2020; 21: 2877-2882.[DOI]
[18] Taguchi F, Kajioka J, Miyamura T. Prevalence rate and age of acquisition of antibodies against JC virus and BK virus in human sera. Microbiol Immunol, 1982; 26: 1057-1064.[DOI]
[19] Elia F, Villani S, Ambrogi F et al. JC virus infection is acquired very early in life: evidence from a longitudinal serological study. J Neurovirol, 2017; 23: 99-105.[DOI]
[20] Agostini HT, Ryschkewitsch CF, Stoner GL. Genotype profile of human polyomavirus JC excreted in urine of immunocompetent individuals. J Clin Microbiol, 1996; 34: 159-164.[DOI]
[21] Jeong BH, Lee KH, Choi EK et al. Genotyping of the JC virus in urine samples of healthy Korean individuals. J Med Virol, 2004; 72: 281-289.[DOI]
[22] Vanchiere JA, Abudayyeh S, Copeland CM et al. Polyomavirus shedding in the stool of healthy adults. J Clin Microbiol, 2009; 47: 2388-2391.[DOI]
[23] Rotondo JC, Candian T, Selvatici R et al. Tracing Males From Different Continents by Genotyping JC Polyomavirus in DNA From Semen Samples. J Cell Physiol, 2017; 232: 982-985.[DOI]
[24] Comar M, Zanotta N, Croci E et al. Association between the JC polyomavirus infection and male infertility. PLoS One, 2012; 7: e42880.[DOI]
[25] Rudick RA, O’Connor PW, Polman CH et al. Assessment of JC virus DNA in blood and urine from natalizumab-treated patients. Ann Neurol, 2010; 68: 304-310.[DOI]
[26] Matos A, Duque V, Luxo C et al. Individuals infected with JC polyomavirus do not present detectable JC virus DNA in oropharyngeal fluids. J Gen Virol, 2012; 93: 692-697.[DOI]
[27] Google. Gemini 3 Flash (Large language model). 2026. Available at:[Web]
[28] Berger JR, Miller CS, Mootoor Y et al. JC virus detection in bodily fluids: clues to transmission. Clin Infect Dis, 2006; 43: e9-e12.[DOI]
[29] Elphick GF, Querbes W, Jordan JA et al. The human polyomavirus, JCV, uses serotonin receptors to infect cells. Science, 2004; 306: 1380-1383.[DOI]
[30] Assetta B, Maginnis MS, Gracia Ahufinger I et al. 5-HT2 receptors facilitate JC polyomavirus entry. J Virol, 2013; 87: 13490-13498.[DOI]
[31] Ricciardiello L, Laghi L, Ramamirtham P et al. JC virus DNA sequences are frequently present in the human upper and lower gastrointestinal tract. Gastroenterology, 2000; 119: 1228-1235.[DOI]
[32] Comerlato J, Souza-Campos F, Souza-Arantes T et al. Distribution and genetic diversity of the human polyomaviruses JC and BK in surface water and sewage treatment plant during 2009 in Porto Alegre, Southern Brazil. Braz J Biol, 2017; 77: 459-468.[DOI]
[33] Levican J, Levican A, Ampuero M et al. JC polyomavirus circulation in one-year surveillance in wastewater in Santiago, Chile. Infect Genet Evol, 2019; 71: 151-158.[DOI]
[34] Prado T, de Castro Bruni A, Barbosa MRF et al. Performance of wastewater reclamation systems in enteric virus removal. Sci Total Environ, 2019; 678: 33-42.[DOI]
[35] Shaheen MNF, Ahmed N, Rady Badr K et al. Detection and quantification of adenovirus, polyomavirus, and papillomavirus in urban sewage. J Water Health, 2024; 22: 401-413.[DOI]
[36] Dias J, Pinto RN, Vieira CB et al. Detection and quantification of human adenovirus (HAdV), JC polyomavirus (JCPyV) and hepatitis A virus (HAV) in recreational waters of Niterói, Rio de Janeiro, Brazil. Mar Pollut Bull, 2018; 133: 240-245.[DOI]
[37] Addetia A, Phung Q, Bradley BT et al. In Vivo Generation of BK and JC Polyomavirus Defective Viral Genomes in Human Urine Samples Associated with Higher Viral Loads. J Virol, 2021; 95: 12.[DOI]
[38] Bofill-Mas S, Girones R. Excretion and transmission of JCV in human populations. J Neurovirol, 2001; 7: 345-349.[DOI]
[39] Mazzoni E, Pellegrinelli E, Mazziotta C et al. Mother-to-child transmission of oncogenic polyomaviruses BKPyV, JCPyV and SV40. J Infect, 2020; 80: 563-570.[DOI]
[40] Oliveira JM, Veiga D, Martins H et al. First detection of JC polyomavirus in vaginal secretions from nonpregnant women: Exploring sexual transmission as a potential route of infection. Int J Infect Dis, 2025; 159: 107992.[DOI]
[41] Haghighi MF, Seyyedi N, Farhadi A et al. Polyomaviruses BK and JC DNA infection in peripheral blood cells from blood donors. Braz J Infect Dis, 2019; 23: 22-26.[DOI]
[42] Frohman EM, Monaco MC, Remington G et al. JC virus in CD34+ and CD19+ cells in patients with multiple sclerosis treated with natalizumab. JAMA Neurol, 2014; 71: 596-602.[DOI]
[43] Bruno G, Miele G, Abbadessa G et al. Atypical progressive multifocal leukoencephalopathy in a kidney transplant recipient with improving symptoms after immunocompetence recovery. Infez Med, 2020; 28: 87-90.
[44] Chandler S, Isbel N. Progressive multifocal leukoencephalopathy 10 years following transplant: 5HT receptor antagonism as an adjunct to immune reconstitution. BMJ Case Rep, 2022; 15: e252284.[DOI]
[45] Garayeva N, Demir E, Dirim AB et al. Expression of JC virus in a kidney transplant recipient with renal cell carcinoma. Nefrologia, 2024; 44: 2.[DOI]
[46] Jackowiak E, Shah N, Chen H et al. A case of immune reconstitution syndrome complicating progressive multifocal leukoencephalopathy after kidney transplant: Clinical, pathological, and radiographic features. Transpl Infect Dis, 2019; 21: e13162.[DOI]
[47] Pjanic M, Aleckovic-Halilovic M, Basic-Jukic N. JC Virus in Kidney Transplant Population: Are We Cautious Enough? J Clin Med, 2024; 13: 2217.[DOI]
[48] Haley SA, O’Hara BA, Nelson CDS et al. Human polyomavirus receptor distribution in brain parenchyma contrasts with receptor distribution in kidney and choroid plexus. Am J Pathol, 2015; 185: 2246-2258.[DOI]
[49] Morris-Love J, Gee GV, O’Hara BA et al. JC Polyomavirus Uses Extracellular Vesicles To Infect Target Cells. mBio, 2019; 10: e00379-19.[DOI]
[50] O’Hara BA, Morris-Love J, Gee GV et al. JC Virus infected choroid plexus epithelial cells produce extracellular vesicles that infect glial cells independently of the virus attachment receptor. PLoS Pathog, 2020; 16: e1008371.[DOI]
[51] Oberholster L, Mathias A, Perriot S et al. Comprehensive proteomic analysis of JC polyomavirus-infected human astrocytes and their extracellular vesicles. Microbiol Spectr, 2023; 11: e0275123.[DOI]
[52] Chen X, Yang F. Classification and Nomenclature of Extracellular Vesicles. In: Wang Q, Zheng L, editors. Extracellular Vesicles. Springer: Singapore, 2024: 3-7.[DOI]
[53] Scribano S, Guerrini M, Arvia R et al. Archetype JC polyomavirus DNA associated with extracellular vesicles circulates in human plasma samples. J Clin Virol, 2020; 128: 104435.[DOI]
[54] Morris-Love J, O’Hara BA, Gee GV et al. Biogenesis of JC polyomavirus associated extracellular vesicles. J Extracell Biol, 2022; 1: e43.[DOI]
[55] Nelson CDS, Carney DW, Derdowski A et al. A retrograde trafficking inhibitor of ricin and Shiga-like toxins inhibits infection of cells by human and monkey polyomaviruses. mBio, 2013; 4: e00729-00713.[DOI]
[56] Treasure T, Nelson CDS. Inhibition of JC polyomavirus infectivity by the retrograde transport inhibitor Retro-2.1. Microbiol Immunol, 2020; 64: 783-791.[DOI]
[57] O’Hara BA, Atwood WJ. Interferon beta1-a and selective anti-5HT(2a) receptor antagonists inhibit infection of human glial cells by JC virus. Virus Res, 2008; 132: 97-103.[DOI]
[58] Nukuzuma S, Nakamichi K, Nukuzuma C et al. Inhibitory effect of serotonin antagonists on JC virus propagation in a carrier culture of human neuroblastoma cells. Microbiol Immunol, 2009; 53: 496-501.[DOI]
[59] Verma S, Cikurel K, Koralnik IJ et al. Mirtazapine in progressive multifocal leukoencephalopathy associated with polycythemia vera. J Infect Dis, 2007; 196: 709-711.[DOI]
[60] Yoshida H, Ohshima K, Toda J et al. Significant improvement following combination treatment with mefloquine and mirtazapine in a patient with progressive multifocal leukoencephalopathy after allogeneic peripheral blood stem cell transplantation. Int J Hematol, 2014; 99: 95-99.[DOI]
[61] Trentalange A, Calcagno A, Ghisetti V et al. Clearance of cerebrospinal fluid JCV DNA with mirtazapine in a patient with progressive multifocal leukoencephalopathy and sarcoidosis. Antivir Ther, 2016; 21: 633-635.[DOI]
[62] Alwehaibi AI, AlJaber MI, Nahrir S. Favorable Response to Mirtazapine in John Cunningham Virus-related Gray Matter Lesion in a Patient with Human Immunodeficiency Virus. Cureus, 2019; 11: e4255.[DOI]
[63] Wang X, Chen J, Gong J et al. JC virus-induced progressive multifocal leukoencephalopathy in a presumably healthy patient. BMC Neurol, 2022; 22: 462.[DOI]
[64] Cifci B, Yildiz Y, Altin E et al. Successful treatment of HIV-associated progressive multifocal leukoencephalopathy (PML) with mirtazapine, mefloquine, and IVIG combination therapy: a case report. J Neurovirol, 2023; 29: 111-115.[DOI]
[65] Dunham SR, Schmidt R, Clifford DB. Treatment of Progressive Multifocal Leukoencephalopathy Using Immune Restoration. Neurotherapeutics, 2020; 17: 955-965.[DOI]
[66] Sidhu N, McCutchan JA. Unmasking of PML by HAART: unusual clinical features and the role of IRIS. J Neuroimmunol, 2010; 219: 100-104.[DOI]
[67] Tawara T, Kai H, Kageyama M et al. A case report of progressive multifocal leukoencephalopathy during steroid treatment for ANCA-associated renal vasculitis. CEN Case Rep, 2020; 9: 354-358.[DOI]
[68] Hentzien M, Guihot A, de Maindreville D et al. Progressive multifocal leukoencephalopathy despite immune recovery in a HIV/HCV co-infected patient. J Neurovirol, 2020; 26: 607-610.[DOI]
[69] Pinnetti C, Cimini E, Vergori A et al. Use of Pembrolizumab for Treatment of Progressive Multifocal Leukoencephalopathy in People Living with HIV. Viruses, 2022; 14: 970.[DOI]
[70] Chatterjee T, Roy M, Lin RC et al. Pembrolizumab for the treatment of Progressive Multifocal Leukoencephalopathy (PML) in a patient with AIDS: A case report and literature review. IDCases, 2022; 28: e01514.[DOI]
[71] Coltart H, El Bouzidi K, Mulka L et al. Marked clinical and radiological improvement following pembrolizumab therapy for HIV-associated PML-IRIS. AIDS, 2023; 37: 199-200.[DOI]
[72] Boumaza X, Bonneau B, Roos-Weil D et al. Progressive Multifocal Leukoencephalopathy Treated by Immune Checkpoint Inhibitors. Ann Neurol, 2023; 93: 257-270.[DOI]
[73] Li C, Wang H, Liu S et al. Successful treatment of HIV-related progressive multifocal leukoencephalopathy and immunological reconstitution inflammatory syndrome with intravenous human immunoglobulin: a case report. AIDS Res Ther, 2025; 22: 89.[DOI]
[74] Balduzzi A, Lucchini G, Hirsch HH et al. Polyomavirus JC-targeted T-cell therapy for progressive multiple leukoencephalopathy in a hematopoietic cell transplantation recipient. Bone Marrow Transplant, 2011; 46: 987-992.[DOI]
[75] Peghin M, Castaldo N, Tascini C et al. Successful JC virus-targeted T-cell therapy for progressive multifocal leukoencephalopathy in a lung transplant recipient. J Heart Lung Transplant, 2022; 41: 991-996.[DOI]
[76] Pocora MM, Bini P, Berzero G et al. Adoptive JC Virus-Specific T Lymphocytes for the Treatment of Progressive Multifocal Leukoencephalopathy: Experience from Two Italian Centers. Viruses, 2025; 17: 934.[DOI]
[77] Möhn N, Grote-Levi L, Bonifacius A et al. Virus-Specific T Cells and Response to Checkpoint Inhibitors in Progressive Multifocal Leukoencephalopathy. JAMA Neurol, 2026; 83: 280-289.[DOI]
[78] Torkildsen Ø, Bru ANS, Behzadi GIN et al. Successful Treatment of Progressive Multifocal Leukoencephalopathy With Tenofovir Alafenamide Fumarate. Neurol Neuroimmunol Neuroinflamm, 2026; 13: e200522.[DOI]
[79] Kaiserman J, O’Hara BA, Haley SA et al. An Elusive Target: Inhibitors of JC Polyomavirus Infection and Their Development as Therapeutics for the Treatment of Progressive Multifocal Leukoencephalopathy. Int J Mol Sci, 2023; 24: 8580.[DOI]
[80] Miyara M, Amoura Z, Parizot C et al. The immune paradox of sarcoidosis and regulatory T cells. J Exp Med, 2006; 203: 359-370.[DOI]
[81] Wills AB, Adjemian J, Fontana JR et al. Sarcoidosis-associated Hospitalizations in the United States, 2002 to 2012. Ann Am Thorac Soc, 2018; 15: 1490-1493.[DOI]
[82] Yoon HY, Kim HM, Kim YJ et al. Prevalence and incidence of sarcoidosis in Korea: a nationwide population-based study. Respir Res, 2018; 19: 158.[DOI]
[83] Guffroy A, Solis M, Gies V et al. Progressive multifocal leukoencephalopathy and sarcoidosis under interleukin 7: The price of healing. Neurol Neuroimmunol Neuroinflamm, 2020; 7: e862.[DOI]
[84] Rosenkranz SC, Häußler V, Kolster M et al. Treating sarcoidosis-associated progressive multifocal leukoencephalopathy with infliximab. Brain Commun, 2022; 4: fcab292.[DOI]
[85] Joy S, Agarwal A, Garg D et al. Sarcoidosis presenting as progressive multifocal leukoencephalopathy in an apparently immunocompetent adult. J Neuroimmunol, 2023; 383: 578201.[DOI]
[86] Chen X, Bäumel M, Männel DN et al. Interaction of TNF with TNF receptor type 2 promotes expansion and function of mouse CD4+CD25+ T regulatory cells. J Immunol, 2007; 179: 154-161.[DOI]
[87] Drew GC, Stevens EJ, King KC. Microbial evolution and transitions along the parasite-mutualist continuum. Nat Rev Microbiol, 2021; 19: 623-638.[DOI]
[88] Casadevall A, Pirofski LA. The damage-response framework of microbial pathogenesis. Nat Rev Microbiol, 2003; 1: 17-24.[DOI]
[89] Tan K, Roda R, Ostrow L et al. PML-IRIS in patients with HIV infection: clinical manifestations and treatment with steroids. Neurology, 2009; 72: 1458-1464.[DOI]
[90] Cobb NA. Contributions to an Economic Knowledge of Australian Rusts (Uredineae).: Improving Wheat by Selection. C. Potter, Government printer, 1894.
[91] Caldwell RM, Schafer JF, Compton LE et al. Tolerance to Cereal Leaf Rusts. Science, 1958; 128: 714-715.[DOI]
[92] Schneider DS, Ayres JS. Two ways to survive infection: what resistance and tolerance can teach us about treating infectious diseases. Nat Rev Immunol, 2008; 8: 889-895.[DOI]
[93] Sanchez KK, Chen GY, Schieber AMP et al. Cooperative Metabolic Adaptations in the Host Can Favor Asymptomatic Infection and Select for Attenuated Virulence in an Enteric Pathogen. Cell, 2018; 175: 146-158.e15.[DOI]
[94] Ahmed W, Wan C, Goonetilleke A et al. Evaluating sewage-associated JCV and BKV polyomaviruses for sourcing human fecal pollution in a coastal river in Southeast Queensland, Australia. J Environ Qual, 2010; 39: 1743-1750.[DOI]
[95] Fumian TM, Guimarães FR, Pereira Vaz BJ et al. Molecular detection, quantification and characterization of human polyomavirus JC from waste water in Rio De Janeiro, Brazil. J Water Health, 2010; 8: 438-445.[DOI]
[96] Fumian TM, Vieira CB, Leite JPG et al. Assessment of burden of virus agents in an urban sewage treatment plant in Rio de Janeiro, Brazil. J Water Health, 2013; 11: 110-119.[DOI]
[97] McQuaig SM, Scott TM, Lukasik JO et al. Quantification of human polyomaviruses JC Virus and BK Virus by TaqMan quantitative PCR and comparison to other water quality indicators in water and fecal samples. Appl Environ Microbiol, 2009; 75: 3379-3388.[DOI]
[98] Schmitz BW, Kitajima M, Campillo ME et al. Virus Reduction during Advanced Bardenpho and Conventional Wastewater Treatment Processes. Environ Sci Technol, 2016; 50: 9524-9532.[DOI]
[99] Torres C, Barrios ME, Cammarata RV et al. High diversity of human polyomaviruses in environmental and clinical samples in Argentina: Detection of JC, BK, Merkel-cell, Malawi, and human 6 and 7 polyomaviruses. Sci Total Environ, 2016; 542: 192-202.[DOI]
[100] Vigiser I, Piura Y, Kolb H et al. JCV seroconversion rate during the SARS COVID-19 pandemic. Mult Scler Relat Disord, 2022; 68: 104244.[DOI]
[101] Krieger SC, Sinks S, Huang F et al. The impact of social distancing measures on anti-JC virus serostatus changes before and during the COVID-19 pandemic in US patients with multiple sclerosis. Mult Scler, 2024; 30: 645-653.[DOI]
[102] Siripurapu R, Ota Y. Human Immunodeficiency Virus: Opportunistic Infections and Beyond. Neuroimaging Clin N Am, 2023; 33: 147-165.[DOI]
[103] Koutsavlis I. Progressive multifocal leukoencephalopathy in multiple myeloma. A literature review and lessons to learn. Ann Hematol, 2021; 100: 1-10.[DOI]
[104] Paolucci M, Gentile L, Gentile M et al. Progressive multifocal leukoencephalopathy in multiple myeloma: a case report of a patient with SARS-CoV-2 infection and an updated systematic literature review. Neurol Sci, 2023; 44: 2995-2998.[DOI]
[105] Pessoa-Gonçalves YM, Farnesi-de-Assunção TS, de Sousa MAD et al. Progressive multifocal leukoencephalopathy triggered by COVID-19 in a previously asymptomatic person living with undiagnosed HIV infection. Int J Infect Dis, 2023; 137: 1-3.[DOI]
[106] Lv Y, Liu X. Hemorrhagic cystitis induced by JC polyomavirus infection following COVID-19: a case report. BMC Urol, 2024; 24: 87.[DOI]
[107] Gosert R, Kardas P, Major EO et al. Rearranged JC virus noncoding control regions found in progressive multifocal leukoencephalopathy patient samples increase virus early gene expression and replication rate. J Virol, 2010; 84: 10448-10456.[DOI]
[108] Iannetta M, Bellizzi A, Lo Menzo S et al. HIV-associated progressive multifocal leukoencephalopathy: longitudinal study of JC virus non-coding control region rearrangements and host immunity. J Neurovirol, 2013; 19: 274-279.[DOI]
[109] McIlroy D, Halary F, Bressollette-Bodin C. Intra-patient viral evolution in polyomavirus-related diseases. Philos Trans R Soc Lond B Biol Sci, 2019; 374: 20180301.[DOI]
[110] Kruzel-Davila E, Divers J, Russell GB et al. JC Viruria Is Associated With Reduced Risk of Diabetic Kidney Disease. J Clin Endocrinol Metab, 2019; 104: 2286-2294.[DOI]
[111] Kondo Y, Windrem MS, Zou L et al. Human glial chimeric mice reveal astrocytic dependence of JC virus infection. J Clin Invest, 2014; 124: 5323-5336.[DOI]
[112] Tan CS, Broge TA, Seung E et al. Detection of JC virus-specific immune responses in a novel humanized mouse model. PLoS One, 2013; 8: e64313.[DOI]
[113] White MK, Gordon J, Berger JR et al. Animal Models for Progressive Multifocal Leukoencephalopathy. J Cell Physiol, 2015; 230: 2869-2874.[DOI]


